The work is based on the following two articles:
• Black Leaf Streak Disease; Pests and Diseases of American Samoa Number 10, American Samoa Community College Community & Natural Resources Cooperative Research & Extension. 2004
• Black Sigatoka An Increasing Thread to Banana Cultivation; Douglas H. Marín, Del Monte Fresh Produce; Ronald A. Romero, Chiquita Brands; Mauricio Guzmán, National Banana Corporation of Costa Rica; Turner B. Sutton, North Carolina State University, Raleigh
Black leaf streak (BLS), or “Black Sigatoka”, is the most damaging leaf disease of bananas worldwide. Most areas of the tropics and subtropics are affected, with reported losses of 30-50%. The first account of BLS in the Pacific was 40 years ago in the Sigatoka Valley, Fiji. Most fungicide used in banana production is to control BLS. The Cavendish-type ‘Williams’ banana, grown by most commercial growers, is very susceptible to the fungal disease. Infected plants have fewer leaves, which leads to fewer and smaller fruits, a delayed harvest, premature ripening, and lower quality fruit.
Potential impact
Black sigatoka causes destruction of banana leaf tissue, which affects the photosynthetic capabilities of the plant and can reduce yields by up to 50%. Black sigatoka is one of the main factors responsible for the decline in banana export industries in South Pacific Nations. Commercial plantations producing bananas for export have to maintain a costly fungicide spray program to control Black sigatoka, and have been criticized on the grounds of environmental and human health considerations. However, if not controlled, fruit produced on diseased plants can ripen prematurely during shipment and cause further losses. The ravages of the disease were controlled by chemical sprays in subsequent years (15-17 fungicide applications) but this considerably increased the cost of production.
Plants affected
Definite symptoms of Black sigatoka have only been recorded on Musa (Bananas), Musa paradisiaca (Plantains), Musa acuminata (Wild banana) and Musa acuminata (subsp. bantesii and subsp. zebrina). Cultivars differ in their reaction to the pathogen. Immunity is not known and it is possible that other wild species and subspecies of Musa are infected, but the disease does not develop significantly. Mycosphaerella fijiensis may also attack the seedling stage of wild bananas.
Season of occurrence
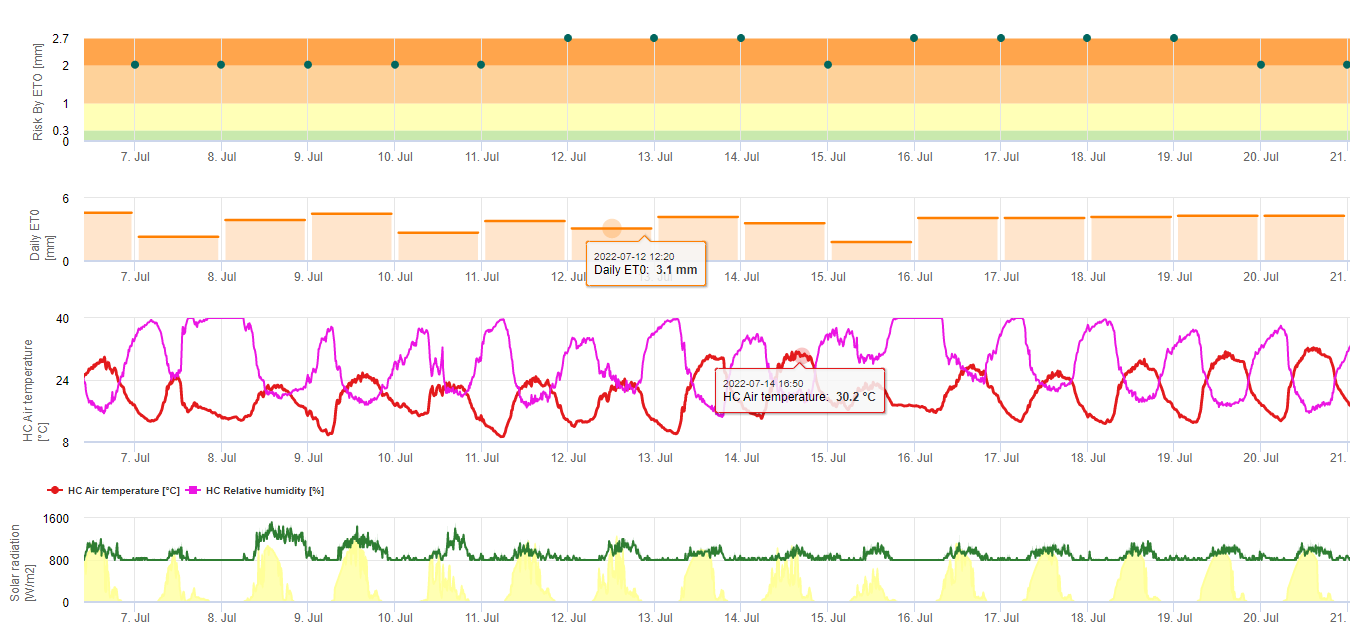
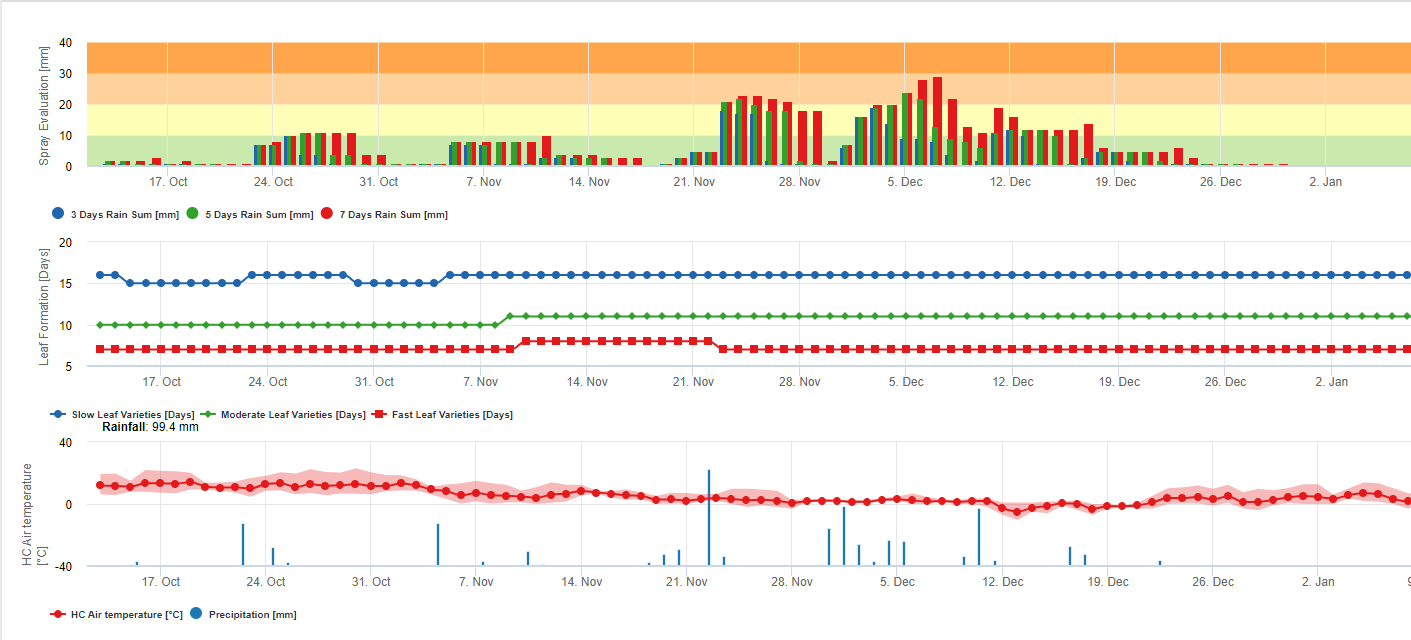
Black sigatoka spores form readily during tropical and sub-tropical summers or under conditions of high humidity and rainfall, especially if there is a film of free water on the leaves. The principle means of spread is through rainwash or splash of spores but later in the development of the disease spores are also forcibly discharged into air currents. Infection occurs on the youngest leaves of the plant during and immediately after unfurling, but older leaves are more conducive to disease development. Obvious visual symptoms appear about 15-20 days after initial infection.
Symptoms
The first apparent symptoms of leaf infection are short, reddish-brown lines about 2 mm long. The lines turn into brown streaks 20-30 mm long which broaden into elliptical spots, brown on the underside of the leaf, black on the upper surface. The spots are slightly sunken and surrounded by a yellowish halo. Next, the center of the large spot becomes gray, bordered by a black line and yellow halo. Finally, the spots merge, killing the leaf.
Epidemiology
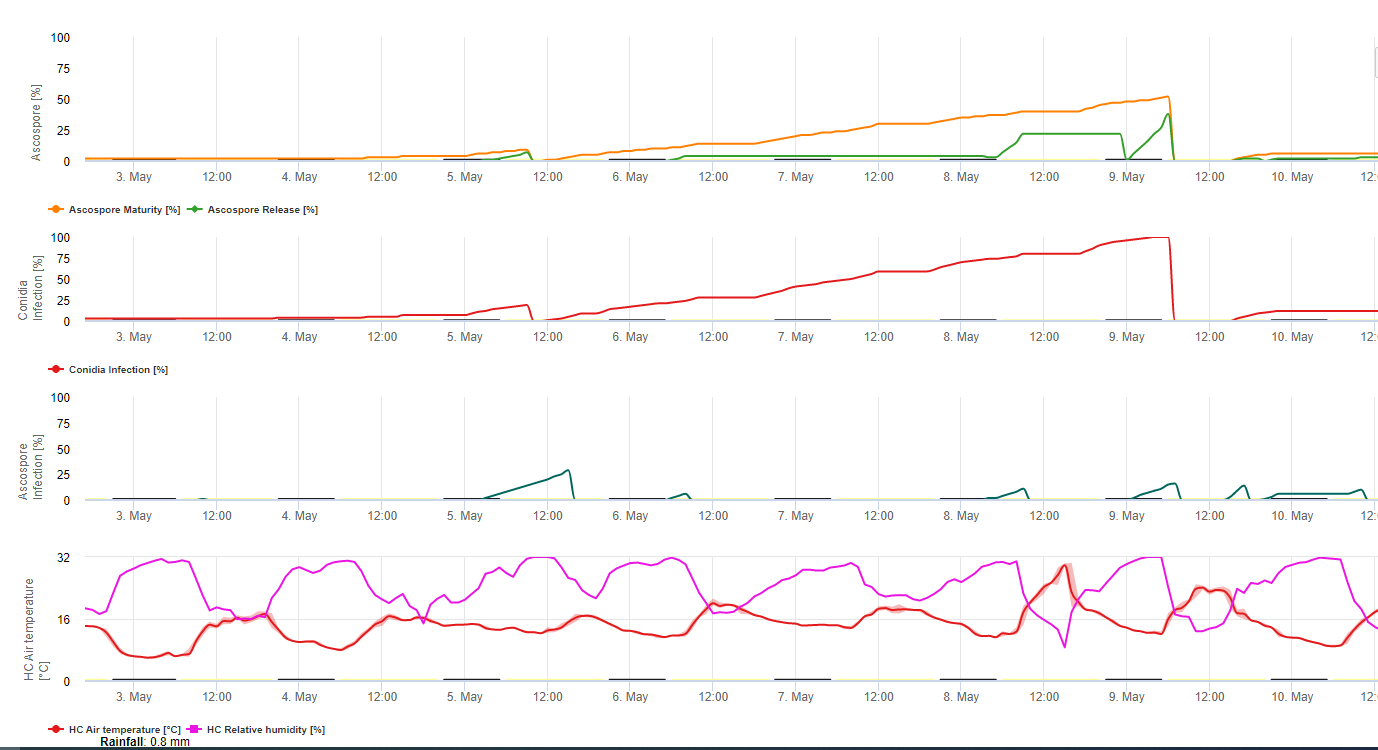
Conidia and ascospores both play roles in the spread of the disease. Conidia form under conditions of high humidity, especially if there is a film of free water on the leaves. They are formed during the development of the first stages of the disease (especially during dash stage 2, streak stage3, and spot stage 4). The principal means of dispersal are rainwash and splash; conidia are not detached by wind. Conidia are associated mostly with local spread of the disease and are important during periods of high humidity, frequent heavy dews, and intermittent showers.
Because M. fijiensis produces relatively few conidia, ascospores are considered to be more important in the spread of black Sigatoka (4,20). However, Jacome et al. (10) stated that conidia are able to cause significant amounts of disease, and the disease symptoms are identical to the ones caused by ascospore infection. He indicated that conidia become more important during dry periods when disease development is delayed because of the presence of less conducive climatic conditions (10). Ascospores are the primary means of dispersal over longer distances within plantations and into new areas, and are the usual means of spread during extended periods of wet weather (3,4,6,10, 11,15).
Ascospores are produced in pseudothecia in mature lesions, which are common on the older leaves of the plant or in dead leaves lying on the ground. Meredith and Lawrence (14) reported that pseudothecia are produced on both sides of the leaf surface, but higher numbers are present on the adaxial surface. Conversely, Gauhl et al. (6) found that more pseudothecia and ascospores were produced on the abaxial leaf surface. Although Burt et al. (2) found that approximately 4.5 ascospores are released per pseudothecium, this number seems very low considering the high concentration of inoculum that is present in the air following rain (4).
Ascospore release
Requires the presence of a film of water from rain or dew that imbibes the pseudothecia and results in the forcible ejection of the ascospores through the leaf boundary layer, where they are disseminated by air currents (15,20). Maturation of pseudothecia requires saturation of the dead leaf tissues for approximately 48 h (3,4,7,10,11,15). Under Hawaiian conditions, ascospore concentrations increased during the night, were highest about 0600 h, and decreased significantly during the day. On rainy days, peak concentrations occurred shortly after rain began. Seasonal increases in daily mean concentrations of ascospores are associated with increased rainfall and relative humidity (15). Gauhl (4), working with bananas, reported similar results under Costa Rican conditions. Whereas Meredith et al. (15) did not find evidence that ascospore production or release was affected by minimum temperatures, Gauhl (4) determined that there is a reduction in the production of inoculum during the drier (or less rainy) months of the year in the Caribbean zone of Costa Rica, which is also the season with the lowest temperatures. Ascospores are dispersed by wind; however, long-distance dispersal is limited to a few hundred kilometers due to their susceptibility to ultraviolet radiation (17). A consistent relationship between ascospore discharge and disease development has not been shown, thus limiting the usefulness of spore trapping for disease forecasting (5,11). Ascospores are deposited mainly on the lower leaf surface during the unfurling of a new leaf, producing a band pattern of infections on the side that is first exposed, which is a reflection of the increased spore deposition on the cylindrical candela leaf during its unfurling as opposed to the entire open lamina of the leaf. Consequently, most infections occur on the abaxial surface of the leaves (3,7).
Infection
Germ tubes take approximately 48 to 72 h to penetrate the stomata (1,20), although the fungus may grow epiphytically on the leaf surface for up to 6 days before actually penetrating the leaves (1). Successful infection is promoted by extended periods of high humidity and the presence of free water on the leaves (3,7,13). Maximum germination occurs when there is free water present. Conidia germinate over a wider range of relative humidity (92 to 100%) than do ascospores (98 to 100%). The effect of temperature on germination can be characterized by a quadratic response function, with an estimated optimum of 26.5°C (10). Stover (21) observed maximum growth of ascospore germ tubes at 26 to 28°C after 24 h incubation. Jacome and Schuh (8,9) reported that older leaves were more susceptible; however, E. Bureau (personal communication) and Romero (18) observed that younger leaves are more susceptible (first to third) than older ones.
Incubation period
The time between infection and the appearance of symptoms varies according to weather conditions and plant susceptibility (68). In bananas, the time of leaf infection is estimated to coincide with the emergence of a new leaf from the apex of the pseudostem (20). Under very favorable conditions in Costa Rica, and with a susceptible host, the incubation period can be as short as 13 to 14 days, whereas during periods of unfavorable weather, the duration of the incubation period can extend up to 35 days. Similar reports exist from Nigeria on plantains (16). During the rainy season, the incubation period was 14 days, but in the dry season it was 24 days. The duration of the incubation period also varies with the susceptibility of the cultivar to M. fijiensis. The incubation period was 26.2 and 25.1 days, respectively, in the cultivars FHIA 1 and FHIA 2, two tetraploid synthetic hybrids developed by FHIA, compared with 22.4 days in the susceptible cultivar Grande Naine (19).
After penetrating the leaf, the hyphae of <M. fijiensis colonize adjacent cells for approximately 7 days without any evidence of disruption of the cells. The vegetative hyphae can emerge from stomata and grow on the surface of the leaf and penetrate adjacent stomata or produce conidiophores and conidia. This epiphytic growth allows the fungus to colonize adjacent leaf tissue, which results in rapid symptom development (4,20). M. fijiensis has a greater ability to penetrate several stomata than does M. musicola, which results in greater spotting than with the former pathogen (20).
Latent period
Although conidia can be produced in lesions exhibiting early symptoms of M. fijiensis and contribute to the epidemic, the latent period is defined by the time the fungus starts to produce lesions with mature pseudothecia and ascospores, which are the main source of inoculum. Like the incubation period, the latent period also varies according to weather conditions, susceptibility of the host, and intensity of infections. The latent period ranged from 25 days during the rainy season (June to December) to 70 days during the dry season at Guapiles, Costa Rica. When the weather is highly conducive for ascospore discharge and infection, many infections occur on the leaves. When infections are dense, they rapidly coalesce at a very early stage of development, accelerating the appearance of mature spots that are characterized by the presence of pseudothecia and ascospores (22). Under these conditions, leaves are rapidly and severely damaged. The latent period also varies according to the level of resistance. For instance, the time from leaf emergence to first mature spot symptom under the same natural conditions for the cultivar Curraré, a cooking banana belonging to the subgroup plantain, was 44 days compared with 34 days for the cultivar Valery, a banana belonging to the subgroup Cavendish (4). The term “disease development time” is widely used in the banana literature in the tropics to refer to the latent period, and it is defined as the time between infection and the formation of mature spots. Another common term used in the banana literature is “symptom evolution time” or “the transition period,” which is the time from first symptoms to the appearance of maturespots (6). The symptom evolution time gives a good indication of how fast the disease is progressing on the leaves.
Literature:
- Beveraggi, A. 1992. Etude des interactions hôte-parasite chez des bananiers sensibles et résistants inoculés par Cercospora fijiensis responsible de la maladie des raies noires. Thèse de 3ème cycle. Université de Montpellier, Montpellier, France.
- Burt, J. P. A., Rosenberg, L. J., Rutter, J., Ramírez, F., and González, O. H. 1999. Forecasting the airborne spread of Mycosphaerella fijiensis, cause of black Sigatoka disease on banana: Estimations of numbers of perithecia and ascospores. Ann. Appl. Biol. 135:369-377.
- Fullerton, R. A. 1994. Sigatoka Leaf Diseases.Pages 12-14 in: Compendium of Tropical Fruit Diseases. R. C. Ploetz, G. A. Zentmyer, W. T. Nishijinia, K. G. Rohrbach, and H. D. Ohr, eds. American Phytopathological Society, St. Paul, MN.
- Gauhl, F. 1994. Epidemiology and Ecology of black Sigatoka (Mycosphaerella fijiensis Morelet) on Plantain and Banana (Musa spp.) in Costa Rica, Central America. INIBAP, Montpellier, France.
- Gauhl, F. 2002. Are spore traps a useful tool to predict Sigatoka disease development in commercial banana plantations. Page 22 in: Int. Workshop Mycosphaerella Leaf Spot Dis. Bananas, 2nd – Programme and Abstracts. (Abstr.)
- Gauhl, F., Pasberg-Gauhl, C., and Jones, D. R. 2000. Disease cycle and epidemiology. Pages 56-62 in: Diseases of Banana, Abacá and Enset. D. R. Jones, ed. CAB International, Wallingford, UK.
- González, M. 1987. Enfermedades del cultivo del banano. Oficina de Publicaciones de la Universidad de Costa Rica, San José.
- Jacome, L. H., and Schuh, W. 1992. Effects of leaf wetness duration and temperature on development of black Sigatoka disease on banana infected by Mycosphaerella fijiensis var. difformis. Phytopathology 82:515-520.
- Jacome, L. H., and Schuh, W. 1993. Effect of temperature on growth and conidial production in vitro, and comparison of infection and aggressiveness in vivo among isolates of Mycosphaerella fijiensis var. difformis. Trop. Agric. 70:51-59.
- Jacome, L. H., Schuh, W., and Stevenson, R. E. 1991. Effect of temperature and relative humidity on germination and germ tube development of Mycosphaerella fijiensis var. difformis. Phytopathology 81:1480-1485.
- Jeger, M. J., Eden-Green, S., Thresh, J. M., Johanson, J. M., Waller, J. M., and Brown, A. E. 1995. Banana diseases. Pages 317-381 in: Bananas and Plantains. S. Gowen, ed. Chapman & Hall, London.
- Marín, D. H., and Romero, R. A. 1992. El combate de la Sigatoka negra. Boletín No. 4, Departamento de Investigaciones, Corporación Bananera Nacional, Costa Rica.
- Mayorga, M. H. 1990. La Raya negra (Mycosphaerella fijiensis Morelet) del plátano y del banano. I. Ciclo de vida del patógeno bajo las condiciones del Urabá. Revista ICA 25:69-77.
- Meredith, D. S., and Lawrence, J. S. 1969. Black leaf streak disease of bananas (Mycosphaerella fijiensis): Symptoms of disease in Hawaii, and notes on the conidial state of the causal fungus. Trans. Br. Mycol. Soc. 52:459-476.
- Meredith, D. S., Lawrence, J. S., and Firman,I. D. 1973. Ascospore release and dispersal on black leaf streak disease of bananas (Mycosphaerella fijiensis). Trans. Br. Mycol. Soc. 60:547-554.
- Mobambo, K. N., Gauhl, F., Pasberg-Gauhl, C., and Zuofa, K. 1996. Season and plant age effect evaluation of plantain for response to black Sigatoka disease. Crop Prot. 15:609- 614.
- Parnell, M., Burt, P. J. A., and Wilson, K. 1998. The influence of exposure to ultraviolet radiation in simulated sunlight on ascospores causing black Sigatoka disease of banana and plantain. Int. J. Biometeorol. 42:22-27.
- Romero, R. A. 1995. Dynamics of fungicide resistant populations of Mycosphaerella fijiensis and epidemiology of black Sigatoka of bananas. Ph.D. diss. North Carolina State, Raleigh.
- Romero, R. A., and Sutton, T. B. 1997. Reaction of four Musa genotypes at three temperatures to isolates of Mycosphaerella fijiensis from different regions. Plant Dis. 81:1139-1142.
- Stover, R. H. 1980. Sigatoka leaf spot of bananas and plantains. Plant Dis. 64:750-756.
- Stover, R. H. 1983. The effect of temperature on ascospore germ tube growth of Mycosphaerella musicola and Mycosphaerela fijiensis var. difformis. Fruits 38:625-628.