
Verificați ce set de senzori este necesar pentru monitorizarea bolilor potențiale ale acestei culturi.

Agenți patogeni
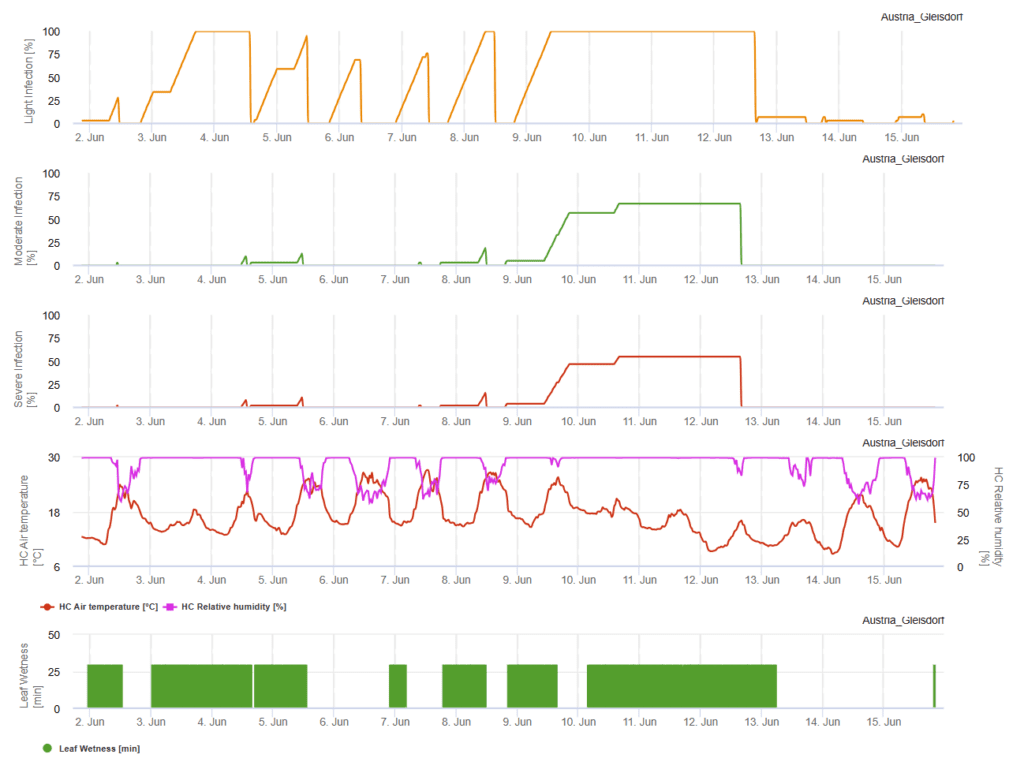
Pyricularia grisea, cunoscut și sub numele de Magnaporthe oryzaeeste o ciupercă patogenă responsabilă de explozia grâului, o boală importantă care afectează culturile de grâu. Agentul patogen se reproduce în principal asexuat prin conidii (spori asexuați), care servesc drept inocul primar pentru inițierea infecțiilor. Aceste conidii sunt produse pe conidiofori și sunt dispersate de vânt, ploaie și mijloace mecanice, cum ar fi echipamentele agricole. La aterizarea pe o gazdă sensibilă de grâu, conidiile germinează și formează appresorii - structuri specializate care facilitează pătrunderea în țesutul vegetal. Odată ajunsă în interior, ciuperca colonizează celulele gazdei, ducând la apariția bolii. Condițiile de mediu joacă un rol crucial în procesul de infectare a P. grisea. Ciuperca se dezvoltă în climatele calde și umede, cu temperaturi cuprinse între 15°C și 30°C și perioade prelungite de umezeală a frunzelor sau umiditate relativă ridicată. Umiditatea continuă a frunzelor timp de cel puțin 14 ore este necesară pentru inițierea infecției, ceea ce face ca acoperirea excesivă de nori, umiditatea ridicată, irigarea excesivă și irigarea intempestivă să favorizeze dezvoltarea bolii. În plus, factorii de stres, cum ar fi seceta, compactarea solului, înălțimile mici de cosire și fertilizarea excesivă cu azot pot exacerba severitatea bolii.
Simptomele
Simptomele inițiale ale exploziei grâului se manifestă prin leziuni înmuiate în apă, în formă de romb, pe frunze, care ulterior devin gri pe măsură ce boala progresează. Aceste leziuni se pot extinde și se pot închega, ducând la arsuri și necroze extensive ale frunzelor. Vârfurile infectate prezintă albire parțială sau completă, pornind adesea de la un punct de infecție gri-negricioasă la rahis sau la baza vârfurilor. În cazul unei presiuni ridicate a inoculului, pot apărea mai multe puncte de infecție pe un singur rahis, ceea ce duce la pierderi semnificative ale randamentului. În plus față de simptomele foliare, P. grisea poate infecta alte părți de deasupra solului ale plantei de grâu, inclusiv nodurile și gâturile, ducând la putrezirea gâtului sau la explozia nodurilor. Acest lucru poate provoca ruperea părților afectate, ceea ce duce la cădere și la reducerea suplimentară a randamentului. Capacitatea agentului patogen de a infecta diferite părți ale plantei de-a lungul sezonului de vegetație îl face deosebit de distructiv, în special în condiții de mediu favorabile.
Modelul FieldClimate
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umezeala frunzelor
Cerul înnorat, condițiile de umiditate și ploile torențiale favorizează dezvoltarea și severitatea bolii. În FieldClimate se calculează trei clase de severitate, iar atunci când se ating 100% de infecție, în câmp au fost create condiții optime pentru agentul patogen fungic. În funcție de soi, trebuie să se ia în considerare măsurile fitosanitare curative din istoricul de teren atunci când o infecție ușoară, moderată sau severă a atins 100%.
Literatură
- Cruz, C. D., & Valent, B. (2017). Boala explozivă a grâului: pericol în mișcare. Patologia plantelor tropicale, 42(3), 210-222.
- Islam, M. T., Croll, D., Gladieux, P., Soanes, D. M., Persoons, A., Bhattacharjee, P., ... & Talbot, N. J. (2016). Apariția exploziei grâului în Bangladesh a fost cauzată de o stirpe sud-americană de Magnaporthe oryzae. BMC Biology, 14(1), 84.
Agenți patogeni
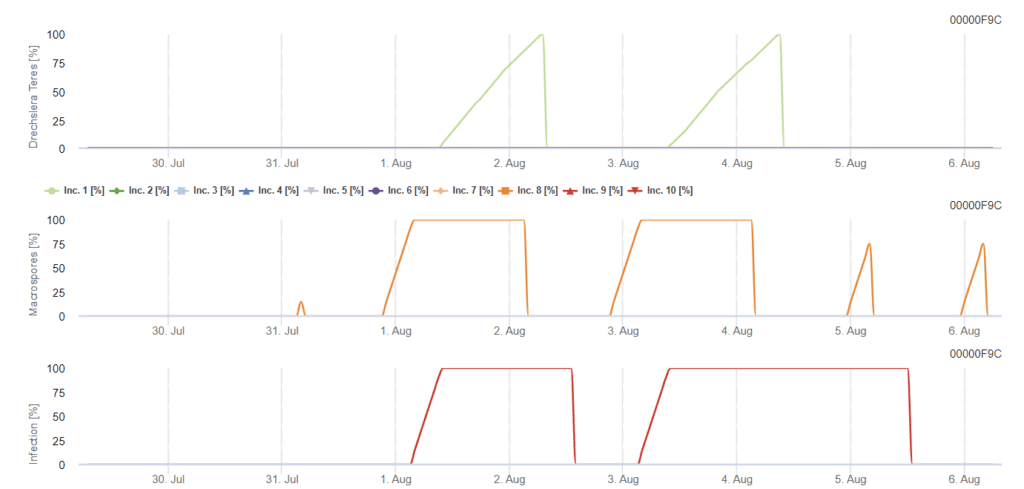
Pyrenophora teres (Drechslera teres) există în două forme - P. teres f. teres și P. teres f. maculata care induc pete în formă de plasă și, respectiv, pete în formă de pată.
Ascosporii eliberați din pseudotecii infectează reziduurile, inițiind infecția primară. În unele cazuri, miceliile și conidiile transmise prin semințe servesc, de asemenea, drept inocul primar. Sporii germinează pe frunze și penetrează peretele celular epidermic exterior, astfel încât agentul patogen se poate dezvolta într-o veziculă intracelulară mare. Urmează stadiul asexuat, în timpul căruia se produc conidii care declanșează infecția secundară, sporind severitatea bolii. La sfârșitul sezonului de vegetație, agentul patogen produce pseudotecii ca sursă de suprasaturare și rămân pe resturile de orz infectat.
Simptomele
Simptomele se manifestă pe frunze, tulpini și boabe, boala progresând de la baza la vârful plantelor. Plantele mai bătrâne au tendința de a suferi daune mai puțin grave, deoarece au o cuticulă mai groasă care împiedică pătrunderea agentului patogen și o capacitate mai mare de a produce compuși antifungici. Agenții patogeni produc toxine care sunt responsabile de simptome, contribuind la necroză și cloroză sau perturbând echilibrul hidric la nivel celular.
În cazul bolii de tip plasă, simptomele apar sub formă de leziuni longitudinale înguste, de culoare maro închis. La soiurile foarte rezistente se vor dezvolta doar câteva leziuni sub formă de puncte mici, fără formarea unui model distinct de tip plasă. În schimb, majoritatea soiurilor sensibile pot prezenta zone clorotice sau îmbibate cu apă în jurul leziunilor necrotice.
Pentru pata de tip pată, simptomele constau în leziuni de culoare maro închis, circulare sau eliptice, care sunt adesea înconjurate de un halou clorotic sau necrotic de lățime variabilă. La soiurile mai puțin sensibile, leziunile sunt în general mai mici și pot fi lipsite de un halou cloros.
Infecțiile severe pot duce la moartea completă a frunzelor, ceea ce le face să pară uscate. De obicei, frunzele mai bătrâne sunt primele care prezintă ofilire, urmate de frunzele mai tinere.
Modelul FieldClimate
Drechslera teres Model
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umezeala frunzelor
- Precipitații
Inoculul primar se dezvoltă la temperaturi optime cuprinse între 15°C și 25°C, cu o temperatură optimă de 20°C. Condițiile de peste 6 ore de întuneric conduc la o creștere optimă a conidiei, așa cum se arată într-un grafic de sporulare a conidiei. Dacă graficul sporulației atinge 100%, condițiile optime sunt stabilite în câmp.
Dispersarea ulterioară are loc prin vânt sau ploaie. Infecțiile țesuturilor vegetale necesită condiții de umiditate, cum ar fi umezeala frunzelor sau o umiditate relativă ridicată timp de aproximativ 10 până la 30 de ore, cu temperaturi optime de 15 °C și 25 °C, după cum se arată într-un grafic al infecției. Atunci când ajunge la 100%, se stabilește că au fost măsurate condițiile optime pentru infecție în câmp. Lumina soarelui poate, de asemenea, favoriza creșterea conidoforilor care produc conidii în urma condițiilor modificate de perioade umede și uscate cu radiații solare ridicate și umiditate relativă scăzută, iar vântul favorizează dezvoltarea bolii.
Literatură
- Backes, A., Guerriero, G., Ait Barka, E., & Jacquard, C. (2021). Pyrenophora teres: taxonomie, morfologie, interacțiune cu orzul și mod de control. Frontiere în știința plantelor, 12, 614951.
- Liu, Z., Ellwood, S. R., Oliver, R. P. și Friesen, T. L. (2011). Pyrenophora teres: profilul unui agent patogen din ce în ce mai dăunător pentru orz. Patologia moleculară a plantelor, 12(1), 1-19.
- Obst, A., & Paul, VH (1993). Boli și dăunători ai cerealelor, Verlag Th. Mann, Gelsenkirchen-Buer.
Agenți patogeni
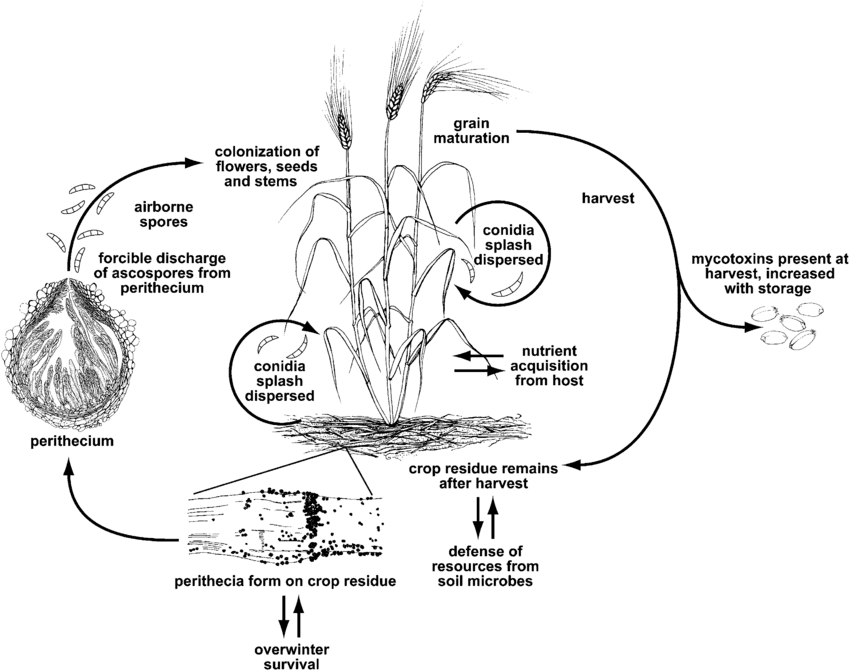
Fusarium head blight sau scab este cauzat de genul Fusarium. Cea mai comună specie este Fusarium graminearum dar speciile cele mai răspândite pot diferi în funcție de regiuni.
F. graminearum poate infecta plantele de grâu în orice stadiu de creștere. Supraviețuiește iarna pe reziduurile de cultură infectate, iar primăvara, infecția primară are loc atunci când ascosporii din peritecii și conidiile sunt dispersate pe glumele sau anterele grâului. Boabele infectate produc boabe zbârcite sau ofilite, adesea numite boabe "de mormânt". Chiar și atunci când boabele par neafectate, ele pot fi contaminate cu micotoxine. Infecția secundară are loc prin conidii, dar epidemiile de grâu depind în primul rând de cantitatea de inocul primar, mai degrabă decât de sursele secundare.
Fusarium head blight conduce, de asemenea, la acumularea de micotoxine dăunătoare în culturi, reprezentând un risc potențial pentru animale. Aceste micotoxine dezactivează mecanismul de apărare al plantelor sau protejează ciuperca împotriva altor organisme. Principala toxină este deoxinivalenolul (DON) și inhibă sinteza proteinelor, perturbând funcționarea normală a celulelor. Oamenii care au consumat grâu cu DON vor prezenta simptome precum greață, febră sau vărsături.
Simptomele

La grâu și la grâul dur, orice parte sau întreaga căpățână poate apărea albită. Căpățânile parțial albe și verzi permit diagnosticarea bolii la grâu. Ciuperca poate infecta și tulpina, provocând o decolorare de la maro la violet. Pe spicele și glumele infectate sunt adesea observate mase de spori de culoare roz până la portocaliu somon în condiții de umiditate. Multe boabe sunt zbârcite și ușoare și sunt uneori numite "pietre funerare" pentru aspectul lor calcaros și lipsit de viață. În condiții favorabile, infecția se poate răspândi la spikele adiacente și poate infecta întregul spike, inclusiv rahisul și pedunculul. Sâmburii pot avea un aspect tern și o decolorare roz, în cazul în care infecția a apărut târziu în faza de dezvoltare.
La orz, spicele infectate se albesc și prezintă un aspect îmbibat cu apă. Boabele pot prezenta, de asemenea, o decolorare și, ocazional, se pot observa mase de spori de culoare somon-portocaliu pe spikelet și glume în condiții de umiditate.
Modele FieldClimate
Estimarea nivelului de FHB într-un câmp se bazează pe numărarea capetelor sau a boabelor afectate. Scala vizuală colorată pentru estimarea severității FHB la grâu este disponibilă la NDSU Extension Service (publicația PP-1095): https://library.ndsu.edu/ir/bitstream/handle/10365/9187/PP1095_1998.pdf?sequence=1&isAllowed=y.
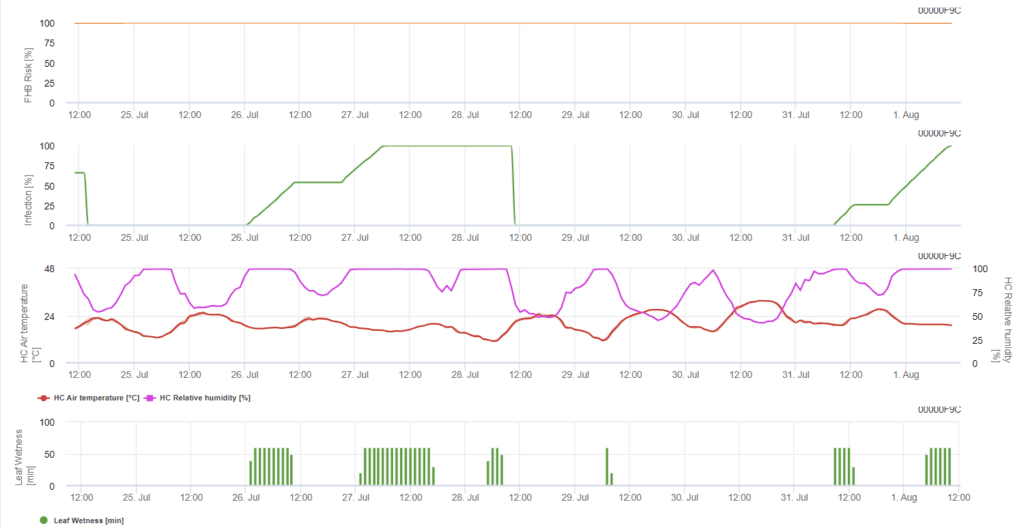
Model de infecție cu Fusarium Head Blight (risc de infecție cu Fusarium Head Blight)
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umiditatea frunzelor
Modelul determină perioadele riscante pentru o infecție. Ori de câte ori se atinge nivelul de infecție 100%, riscul este foarte ridicat, iar condițiile pentru ciupercă au fost favorabile pentru infecție. Cunoștințele fermierului cu privire la stadiul de dezvoltare al diferitelor soiuri de grâu oferă posibilitatea de a decide aplicarea unui spray curativ imediat după infecție.
Agenții fungici ai Fusarium Head Blight la grâu sunt favorizați de temperaturile calde de 20-30 °C și de perioadele lungi de umiditate. Perioadele de umezeală a frunzelor de câteva zile vor duce la apariția simptomelor vizibile timpurii. Simptomele pot fi observate și după o lungă perioadă de latență, dacă infecția este urmată de o perioadă de umezeală a frunzelor de 18 ore sau chiar mai scurtă și după un inocul artificial la temperaturi de 15°C.
Rezumând toate combinațiile diferite de temperatură și umiditate pe care le-am găsit în literatura de specialitate, am decis să semnalăm infecțiile cu Fusarium Head Blight dacă temperatura și perioada sau perioadele de umezeală foliară cu o umiditate relativă mai mare de 85% depășesc valorile vizualizate în graficul următor. Calculul valorii progresului urmează relația dintre durata condițiilor de umezeală și temperatură.
Model de alertă privind micotoxina Fusarium (Fusarium Head Blight)
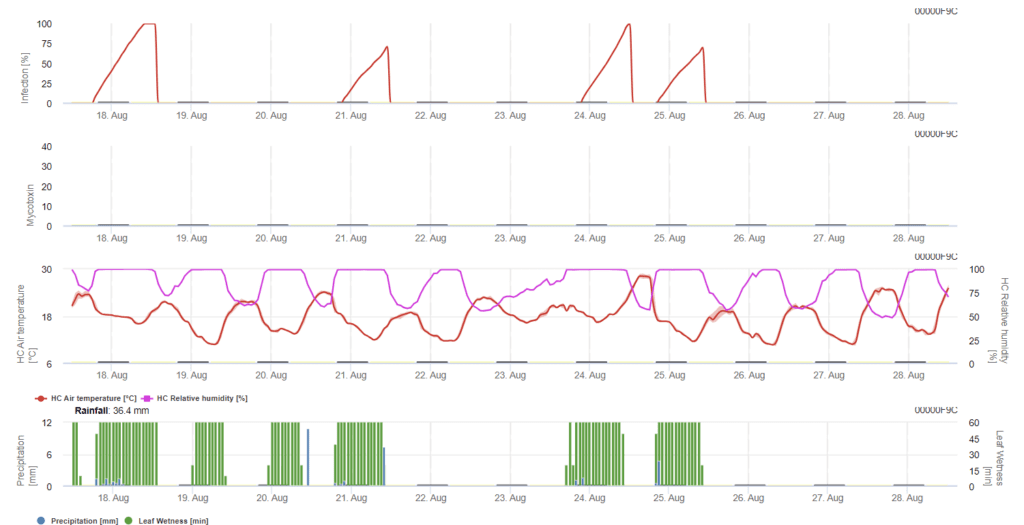
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umiditatea frunzelor
- Precipitații
în acest model, infecția cu FHB este calculată în funcție de precipitații (2 mm necesari), umiditate relativă (peste 85%) sau umezeală foliară, temperatură în timpul procesului. În cazul în care infecția atinge 100%, condițiile optime pentru ciuperca patogenă au fost atinse. În continuare, modelul calculează riscul de micotoxine FHB.
Testele de infectare cu perioade îndelungate de umezeală a frunzelor cu Fusarium head blight conduc la un conținut ridicat de micotoxine. Pe baza acestor informații, se presupune că o perioadă de umezeală a frunzelor de 48 de ore sau mai mult în timpul etapelor 61 și 69 prezintă un risc ridicat de micotoxine.
Experiențele acumulate pe parcursul analizei DON în grâul cultivat comercial au arătat că perioadele de umezeală a frunzelor suficient de lungi pentru infecție după o infecție inițială în stadiile 61-69 pot crește valorile DON. În cazul unor perioade mai lungi de umezire a frunzelor, micotoxinele pot crește până la stadiul 85.
- Stadiul BBCH 61: Începutul înfloririi; primele antere vizibile
- Stadiul BBCH 69: Sfârșitul înfloririi; toate spicele au terminat înflorirea, dar pot rămâne unele antere deshidratate
- Stadiul BBCH 85: În mijlocul coacerii; aluat moale cu conținut de boabe moale, dar uscat, impresia unghiei nu este menținută
Modelul acumulează cifra riscului proporțional cu progresul infecției pentru fiecare perioadă de infecție reușită din perioada selectată pentru calcul. Șase infecții finalizate ar conduce la un risc de 100%. În mod normal, perioada de umezeală a frunzelor care conduce la o infecție cu fusarium este mai lungă decât minimul necesar. Prin urmare, majoritatea infecțiilor cu fusarium vor duce la o creștere a riscului mai mare de 17%.
Valoarea riscului de micotoxine pe istoricul câmpului. Grâul cultivat după nefolosirea solului poate prezenta un risc mic doar dacă nu este pulverizat în situația optimă. În cazul grâului neprotejat, trebuie să ne așteptăm la o creștere a valorilor DON după 35% a riscului. Grâul după grâul fără arat după orice altă cultură poate prezenta un risc mai mare de 50%. Dacă avem grâu după grâu cu lucrare a solului, riscul poate fi crescut până la 70%. Grâul din primul an trebuie testat pentru DON dacă riscul ajunge la 100%.
Literatură
- https://www.fao.org/4/y4011e/y4011e0j.htm
- https://scabusa.org/pdfs/NDSU_PP-804_FHB-Small-Grains.pdf
- Lancashire, P. D., Bleiholder, H., Boom, T. V. D., Langelüddeke, P., Stauss, R., Weber, E., & Witzenberger, A. (1991). Un cod zecimal uniform pentru stadiile de creștere ale culturilor și buruienilor. Analele de biologie aplicată, 119(3), 561-601.
- Trail, F. (2009). Pentru valuri de grâne bolnave: Fusarium graminearum în era postgenomică. Fiziologia plantelor, 149(1), 103-110.
- Schumann, G. L. (2010). Instructor în domeniul sănătății plantelor| Volume: 10| Year: 2010| Article Type: Lesson Plans. Sănătatea plantelor, 10.
Agenți patogeni
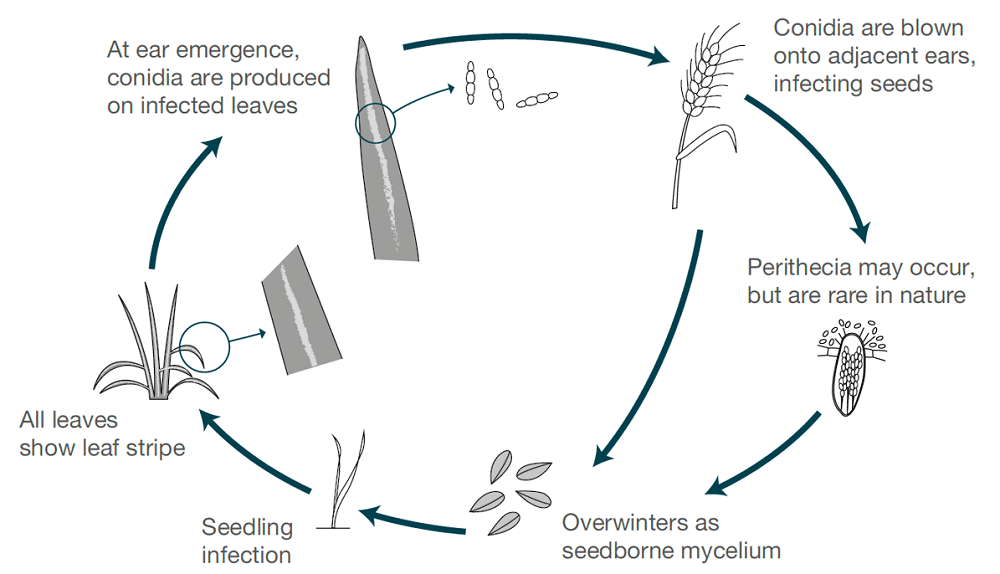
Zgârietura orzului este cauzată de Pyrenophora graminea (Drechslera graminea), un agent patogen transmis prin semințe care supraviețuiește în epiderma semințelor infectate. Ciuperca infectează răsadurile în condiții reci și umede. Ciuperca se dezvoltă sistemic în interiorul plantei, infectează întreaga plantă, produce toxine, ucide celulele și decolorează țesutul foliar între nervuri, provocând astfel leziuni striate. Dungile sunt mai frecvente la orzul de iarnă decât la orzul de primăvară.
Când condițiile sunt umede sau umede, sporii sunt produși pe suprafața frunzelor atunci când înfloresc vârfurile sănătoase ale plantelor. Sporii sunt împrăștiați de vânt către aceste vârfuri în dezvoltare, germinează și provoacă infecții. Boabele sunt cele mai sensibile în timpul dezvoltării timpurii. În fiecare sezon are loc un singur ciclu de infecție și de producere a sporilor.
Simptomele

Simptomele sunt mai ales proeminente în stadiul de apariție a urechii.
Dungile de la verde pal la galben apar inițial în partea bazală a frunzei și a tecii frunzei. Aceste dungi devin treptat maro până la maro închis, urmate de uscarea și despicarea limbului frunzei. Acestea se extind pe întreaga lungime a frunzei, devin necrotice și se unesc, ducând în cele din urmă la moartea plantei.
Plantele pot părea pipernicite, pot produce puțini taleri și, în cazurile grave, nu produc căpățâni sau semințe. Lungimea spicului poate fi, de asemenea, redusă din cauza boabelor brune slab dezvoltate.
Modelul FieldClimate
Pyrenophora graminea Model
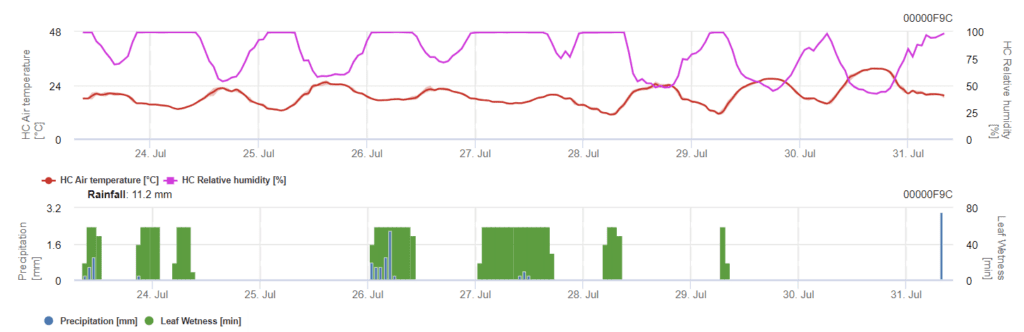
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umiditatea frunzelor
- Precipitații
Modelul prezintă două etape. Prima etapă este reprezentată de perioadele riscante din timpul perioadelor de urgență ale plantei, stadiile incipiente. A doua etapă este în timpul perioadei de înflorire și de dezvoltare a căpățânii de orz. Atunci când infecția ajunge la 100%, în câmp au fost create condițiile optime pentru ca ciuperca să infecteze țesuturile plantei și să se dezvolte sistematic în interior.
Literatură
- Richardson, M. J. (1996). Micologia semințelor. Cercetări micologice, 100(4), 385-392. https://ahdb.org.uk/knowledge-library/barley-leaf-stripe-life-cycle-and-disease-symptoms https://plantwiseplusknowledgebank.org/doi/full/10.1079/pwkb.species.46115
Agenți patogeni
Există două boli Septoria majore la grâu - Septoria tritici blotch cauzată de Septoria tritici și Septoria nodorum blotch cauzate de Septoria nodorum.
Septoria tritici supraviețuiește pe reziduurile de cultură infectate în timpul verii și începe să infecteze plantele de grâu în toamnă. Ciuperca se dezvoltă în condiții reci și umede. Are două etape - o fază latentă și o fază necrotică. În timpul fazei latente, ciuperca inoculează țesuturile vegetale noi și, pătrunzând prin stomate, urmează colonizarea, pe care o dezvoltă intercelular în țesuturile vegetale. S. tritici nu necesită structuri fizice de hrănire, ci se hrănește prin extragerea nutrienților din celulele gazdă care și-au pierdut integritatea structurală. Plantele pot prezenta un răspuns hipersensibil și se pot dezvolta leziuni necrotice pe țesuturile afectate ca răspuns la infecție.
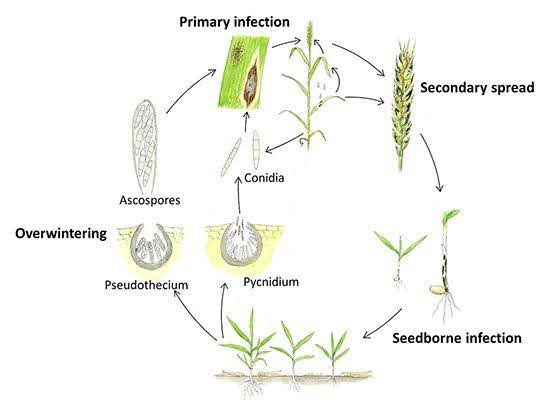
Septoria nodorum supraviețuiește pe parcursul verii sub formă de micelii, picnidii și pseudotecii în stare latentă pe reziduurile de cultură. Infecția începe de obicei toamna, când pycnidiosporele din pycnidia și ascosporele din pseudotecia sunt dispersate de vânt și ploaie, inițiind infecția primară. Acești spori pătrund în cuticula frunzei și formarea picnidiei urmează după infecție. Infecția secundară are loc pe măsură ce se produc și se dispersează pycnidiospori de la frunzele inferioare la frunzele superioare și glume. Agentul patogen produce, de asemenea, toxine care contribuie la dezvoltarea bolii.
Simptomele
Septoria tritici blotch
În toamnă, Septoria tritici simptomele apar inițial ca mici pete galbene pe frunze, care conțin mici pete negre, care sunt corpuri fructifere fungice. Leziunile au o formă neregulată, de la eliptice la lungi și înguste, se extind și devin maro până la maro roșiatic la maturitate. Boala debutează de obicei pe frunzele inferioare și se deplasează treptat în sus, afectând în cele din urmă frunza steag. În condiții de umezeală, ciuperca se poate răspândi la căpățânile de grâu, provocând leziuni brune pe glume și coame, cunoscute sub denumirea de "glume blotch". Septoria tritici blotch poate fi confundată cu alte boli ale grâului. Cu toate acestea, prezența corpurilor fungice negre este o caracteristică esențială a S. tritici. Deși Septoria nodorum poate produce, de asemenea, corpuri fructifere maro în condiții umede, acestea diferă în ceea ce privește culoarea și dimensiunea, cu Septoria tritici corpurile fructifere fiind mai mari.
Septoria nodorum blotch
Septoria nodorum simptomele apar mai întâi pe frunzele superioare. Inițial, pe frunze apar pete maro închis cu un halo galben. Arderea vârfului frunzelor este un alt indiciu inițial. Leziunile se extind și devin necrotice cu un centru maro închis pe măsură ce se maturizează. În interiorul leziunilor sunt vizibile mici structuri maro închis, care sunt corpuri fructifere. Conidiile eliberate din corpurile de fructificare au aspectul unor mase de culoare albă până la roz. Frunzele grav infectate pot fi complet acoperite de leziuni, ducând în cele din urmă la moartea țesutului foliar. Ciuperca poate afecta, de asemenea, glumele și coamele: apar leziuni brune similare și boala progresează în jos.
Modelul FieldClimate
Infecțiile cu Septoria sunt posibile la temperaturi scăzute, în timp ce temperaturile sub 7°C pot să nu conducă la o infecție în decurs de 2 zile. Temperatura optimă a bolii este atinsă în zona de 16-21°C. Infecțiile sunt posibile într-o perioadă de umiditate relativă ridicată sau de umezeală a frunzelor de 14 ore sau mai mult.
Septoria tritici Model
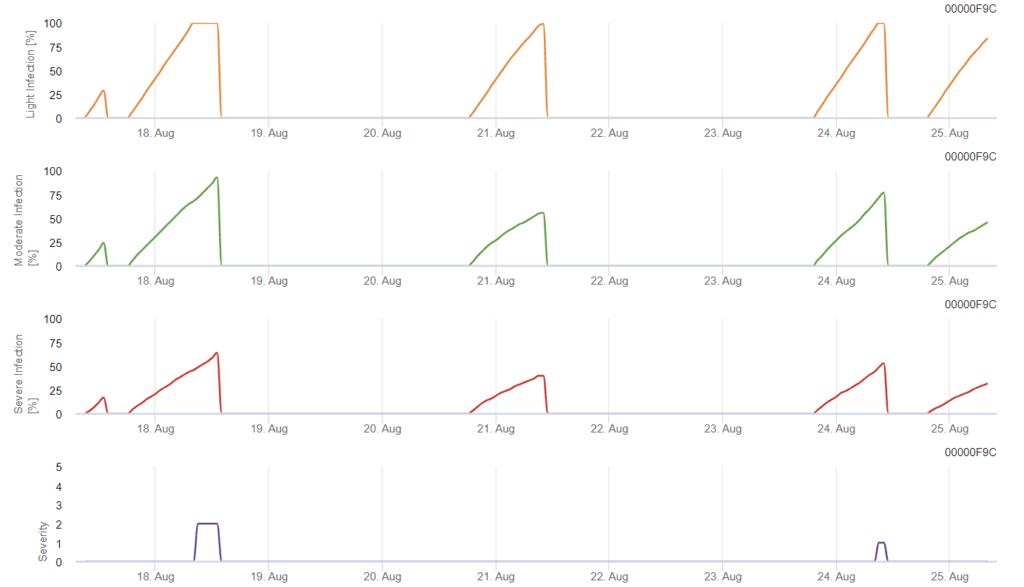
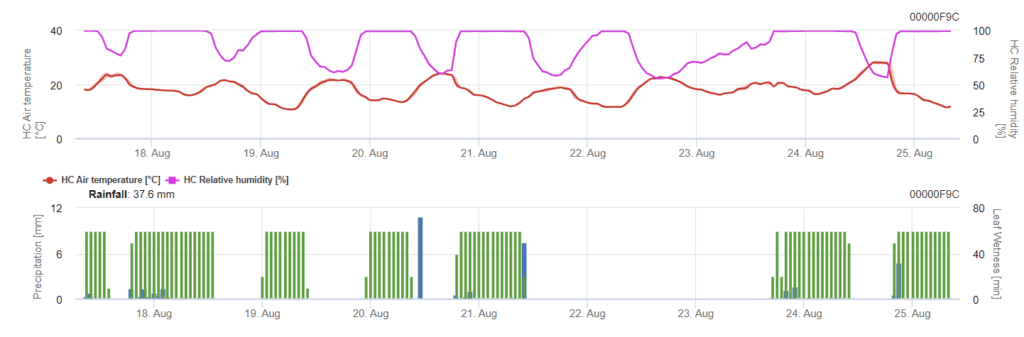
Infecția începe după o ploaie de 0,5 mm. Temperaturile optime pentru infecțiile cu Septoria sunt în jur de 16°C până la 21°C și umezeala frunzelor sau umiditatea relativă ridicată timp de 14 ore. Septoria tritici este calculată în trei clase de severitate. În plus, determinăm niveluri de severitate care sprijină evaluarea presiunii infecției între stadiile BBCH 10-32 ale grâului.
- Stadiul BBCH 10: Dezvoltarea frunzei; prima frunză prin coleoptil
- Stadiul BBCH 32: Alungirea tulpinii; nodul 2 la cel puțin 2 cm deasupra nodului 1
- Stadiul BBCH 51: Apariția și răsărirea inflorescenței; începutul răsăririi cu vârful inflorescenței ieșit din teacă și primul spic abia vizibil.
Am decis să nu utilizăm un model pentru formarea picnidiei. Condiția necesară pentru formarea pycnidiei se presupune a fi o perioadă cu umiditate relativă mai mare de 85%. Durata de viață a pycnidiei este de 24 de ore. În toate climatele în care ciuperca are șansa de a infecta, vom găsi două ore care îndeplinesc această condiție aproape în fiecare zi în jurul răsăritului soarelui.
Evaluarea severității infecției
Pentru a evalua Septoria tritici presiunea infecției între stadiile 10 și 32 și între 32 și 51, trebuie să evaluăm gravitatea infecțiilor în funcție de condițiile climatice. Severitatea infecției este evaluată pe o scară de la 1 la 5:
- 1: infecție slabă cu ploi mai mici de 5 mm
- 2: infecție slabă cu ploaie mai mare de 5 mm
- 3: infecție moderată cu ploaie mai mică de 5 mm
- 4: infecție moderată cu ploaie mai mare de 5 mm/ infecție severă cu ploaie mai mică de 5 mm
- 5: infecție severă cu ploaie mai mare de 5 mm
Evaluarea presiunii bolii
Există trei factori pentru presiunea bolii în câmp - clima, istoricul câmpului și susceptibilitatea soiului cultivat. Dacă putem acumula valorile severității bolii de la stadiul 10 la stadiul 32 până la valoarea 4, ne putem aștepta la o presiune slabă a bolii din cauza climei. Dacă această valoare ajunge la 6, ne putem aștepta la o presiune moderată a bolii, iar dacă ajunge la 10, la o presiune mai mare a bolii din partea climei.
Cunoașterea susceptibilității soiului și a istoricului câmpului ne va determina să pulverizăm sau nu în cazul unei presiuni slabe sau moderate a bolii în această situație. O valoare acumulată de 10 poate conduce oricum la o pulverizare în stadiul 32.
Decizia de a pulveriza într-un stadiu ulterior depinde de climatul de primăvară. Dacă putem acumula valorile de severitate din stadiul 10 la o valoare de 6, ne putem aștepta la o presiune slabă a bolii. Dacă această valoare ajunge la 10, ne putem aștepta la o presiune moderată a bolii, iar dacă această valoare ajunge la 15, ne putem aștepta la o presiune ridicată a bolii din cauza situației climatice.
În FieldClimate, dispozitivul Septoria tritici împreună cu cele trei severități diferite ale infecției. Datorită precipitațiilor și a perioadelor lungi de umezeală a frunzelor, au fost îndeplinite condițiile pentru o infecție gravă cu S. tritici. Nivelurile de severitate ating cea mai mare valoare de 5, ceea ce înseamnă că există un risc ridicat de infecție.
Septoria nodorum Model
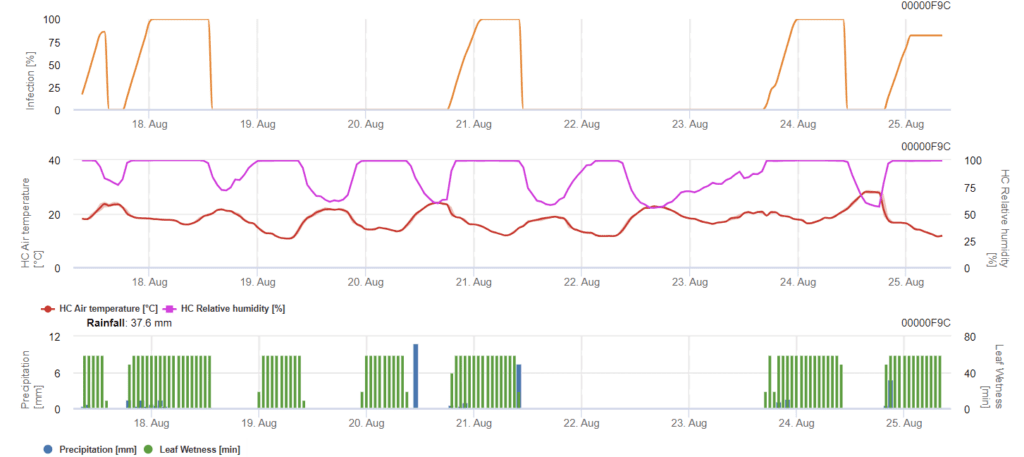
Septoria nodorumbiologia infecției diferă într-o oarecare măsură de Septoria tritici dar diferența nu este suficient de mare pentru un model separat. Prin urmare, sugerăm utilizarea modelului tritici pentru întregul complex al bolii Septoria. În zonele cu presiune ridicată de S. nodorum, infecțiile clasificate ca fiind slabe, cu o valoare a severității de 2, ar trebui tratate mai serios decât în alte domenii.
The Septoria nodorum calculează riscul de apariție a bolii. Măsurile de protecție a plantelor ar trebui luate în considerare dacă riscul atinge 80%. Atunci când riscul este de 100% și infecția este deja determinată, trebuie luate măsuri sistemice de protecție a plantelor (aplicații curative).
Literatură
- Brennan, C. J., Benbow, H. R., Mullins, E. și Doohan, F. M. (2019). O revizuire a necunoscutelor cunoscute în stadiile incipiente ale bolii septoria tritici blotch a grâului. Patologia plantelor, 68(8), 1427-1438.
- De Wolf, E. D. (2008). Septoria tritici blotch. Mehra, L. K., Adhikari, U., Ojiambo, P. S. și Cowger, C. (2019). Septoria nodorum blotch de grâu. Instructorul pentru sănătatea plantelor.
- Solomon, P. S., Lowe, R. G., TAN, K. C., Waters, O. D., & Oliver, R. P. (2006). Stagonospora nodorum: cauza pătării cu stagonospora nodorum a grâului. Patologia moleculară a plantelor, 7(3), 147-156.
Agenți patogeni
Boala frunzelor este cauzată de Rynchosporium secalis.
Infecția primară are loc prin ascospori sau conidii pe resturile de recoltă. Acești spori sunt dispersați de vânt și ploaie, germinează și, pe măsură ce celulele epidermice și mezofile se prăbușesc după o infecție, simptomele devin vizibile. Infecția secundară este provocată de conidiile de pe frunzele infectate. În condiții de umiditate, conidiile germinează pe suprafața frunzei și produc hife care penetrează cuticula direct deasupra celulelor epidermice. Creșterea fungică ulterioară este limitată la regiunea subcuticulară a epidermei.
Simptomele
R. secalis poate infecta orice parte a frunzelor. Petele de formă neregulată sunt principala caracteristică, iar infecția începe de la partea inferioară până la partea superioară a frunzelor. Petele apar inițial ca zone îmbibate cu apă. Pe măsură ce boala progresează, acestea devin cenușii în centru din cauza formării sporilor, cu o margine maro. Pot urma necroza și cloroza. Petele se unesc, distrugând întreaga frunză.
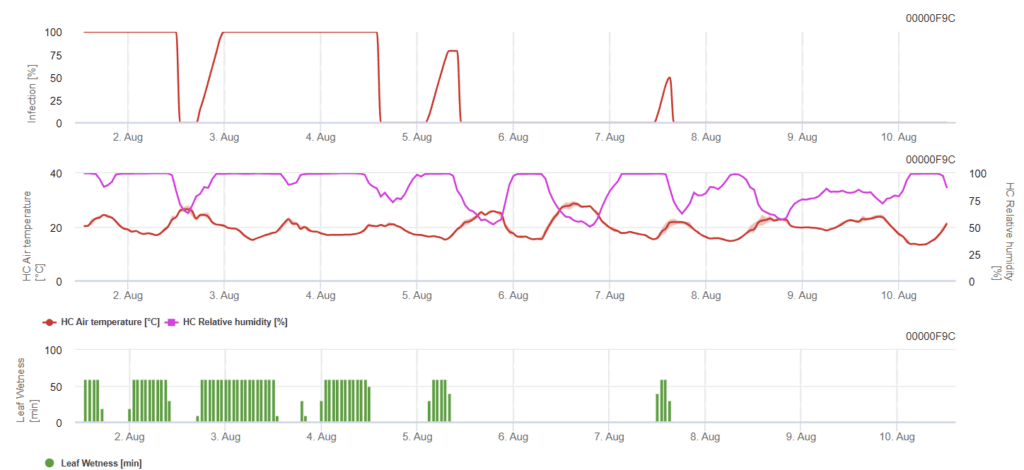
Modelul FieldClimate
Modelul Rynchosporium
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umiditatea frunzelor
Pentru ca o infecție să apară, este nevoie de un minim de 7 până la 15 ore de umezeală a plumbului, în funcție de temperatură (optim la 20'C). Atunci când graficul ajunge la 100%, înseamnă că au fost determinate condițiile optime pe teren.
Literatură
- Brooks, F. T. (1928). Observații privind Rhynchosporium secalis (Oud.) Davis, pătarea frunzelor de orz și secară. New Phytologist, 27(4), 215-219.
- Fowler, A. M., & Owen, H. (1971). Studii privind pătarea frunzelor de orz (Rhynchosporium secalis). Transactions of the British Mycological Society, 56(1), 137-152.
- Zhan, J., Fitt, B. D., Pinnschmidt, H. O., Oxley, S. J. P. și Newton, A. C. (2008). Rezistența, epidemiologia și gestionarea durabilă a populațiilor de Rhynchosporium secalis pe orz. Patologia plantelor, 57(1), 1-14.
Agenți patogeni
Ramularia collo-cygni, agentul cauzal al petei Ramularia la orz, se răspândește în principal prin reproducere asexuată prin conidii, care necesită umiditate pentru germinare și dezvoltare timpurie. După germinarea pe suprafața frunzei, ciuperca pătrunde în frunză prin stomate și colonizează țesutul intercelular. Plantele de orz infectate pot să nu prezinte inițial niciun simptom, dar daunele apar de obicei după înflorire.
Ciclul de viață al R. collo-cygni nu este pe deplin înțeleasă, dar implică transmiterea verticală prin semințe infectate, permițând agentului patogen să supraviețuiască între sezoanele de creștere. Conidiile din aer contribuie, de asemenea, la răspândirea secundară, inițiind noi infecții în condiții favorabile. În timp ce semințele reprezintă principalul mecanism de iernare, rolul gazdelor alternative și al unei structuri fungice secundare denumită Asteromella rămâne incertă și este încă în curs de studiu.
Simptomele

Simptomele apar pe ambele părți ale frunzelor după înflorire în general, dar sunt mai frecvente pe frunzele superioare. Simptomele inițiale apar sub forma unor pete galbene până la maro pe nervurile frunzelor, înconjurate de un halou clorotic. Odată ce apar petele, frunza devine clorotică și necrotică, începând de obicei de la vârful și marginile frunzei. Petele învecinate se pot întâlni, creând zone întunecate mai mari. Pot fi observate și pete punctiforme mici. Deși simptomele petelor de frunze de Ramularia pot fi confundate cu alte boli, caracteristica principală este că petele de frunze sunt delimitate de nervurile frunzelor.
Există așa-numitele 5R pentru a distinge petele de frunze ale Ramularia:
- Inelat cu o margine galbenă de cloroză
- Formă dreptunghiulară
- Limitat de nervurile frunzei
- Colorație roșiatică-maronie
- Chiar prin frunză
Model FieldClimate
Modelul Ramularia
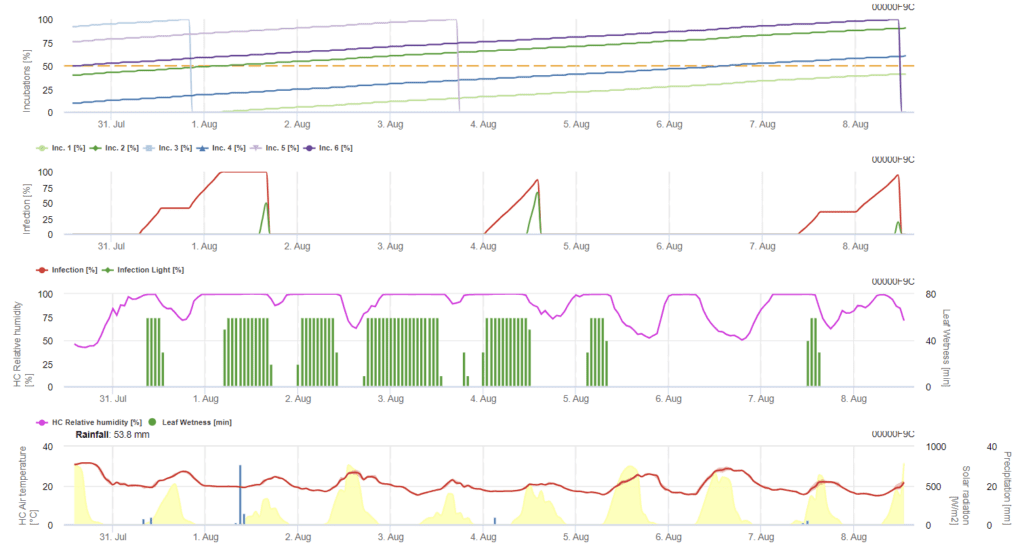
Infecția cu Ramularia pe orz este determinată în principal de condițiile de umiditate. Astfel, începem calculul cu un eveniment de ploaie, urmat de umezeală foliară sau umiditate relativă mai mare de 85%.
Există două moduri de a descrie o infecție:
- Infecție: o dată când condițiile umede doar favorizează o infecție
- Infecție: o dată când este necesară radiația globală pentru a termina un eveniment de infecție
Mai departe, ori de câte ori se atinge infecția 100%, modelul începe să determine timpul de incubare. Atunci când incubarea ajunge la 50%, se recomandă controlul câmpului pentru noi inoculi, iar atunci când ajunge la 100%, se prevăd leziuni necrotice pe frunze.
Literatură
- Havis, N. D., Brown, J. K., Clemente, G., Frei, P., Jedryczka, M., Kaczmarek, J., ... & Hess, M. (2015). Ramularia collo-cygni - un patogen emergent al culturilor de orz. Fitopatologie, 105(7), 895-904.
- Huss, H. (2002, aprilie). Biologia Ramularia collo-cygni. În Proc. Second Int. Workshop Barley Leaf Blights. Aleppo, Siria (pp. 321-328).
- Walters, D. R., Havis, N. D. și Oxley, S. J. (2008). Ramularia collo-cygni: biologia unui patogen emergent al orzului. FEMS Microbiology Letters, 279(1), 1-7. https://grdc.com.au/__data/assets/pdf_file/0025/443509/GRDC_FS2103_Ramularia_03.pdf
Agenți patogeni
Putregaiul rădăcinilor Rhizoctonia la grâu este cauzat de Rhizoctonia solani. Agentul patogen poate supraviețui mulți ani în sol sub formă de scleroți. În unele cazuri, supraviețuiește și sub formă de miceliu pe resturile vegetale. Odată ce temperatura crește, scleroții devin activi și generează o masă de hife, permițând infectarea prin atașarea la rădăcini. Miceliul produce așa-numitele "perne de infecție", iar ciuperca invadează și colonizează țesutul radicular. Infecția rădăcinilor slăbește răsadurile, ceea ce duce la o creștere slabă, și reduce capacitatea de a transporta apă și substanțe nutritive către țesuturile superioare ale plantelor, ducând în cele din urmă la moartea plantelor.
Simptomele
La început, apar pete goale, cu diametrul variind de la câțiva centimetri la câțiva metri. Aceste pete goale sunt adesea asociate cu plante pipernicite care pot prezenta îngălbenire, ofilire sau chiar o colorație purpurie. Plantele infectate pot fi foarte pipernicite și pot prezenta semne de stres cauzat de secetă sau de deficit de nutrienți, ceea ce poate duce, în unele cazuri, la o moarte prematură.
Leziunile brun-roșcate se dezvoltă pe tulpini și rădăcini sub linia solului, iar aceste leziuni se adâncesc pe măsură ce boala progresează. Putregaiul rădăcinilor poate reduce nodularea și face ca cortexul rădăcinii să se rupă ușor, rezultând vârfuri maronii ale lăstarilor.
Modelul FieldClimate
Model de risc pentru Rhizoctonia solani
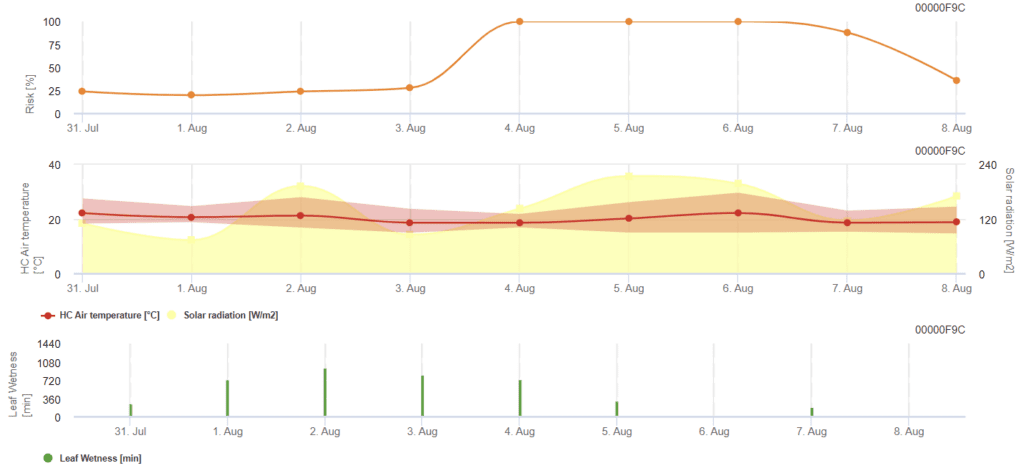
Senzori necesari:
- Temperatura aerului
- Radiația solară
- Umiditatea frunzelor
Modelul calculează perioadele riscante de Rhizoctonia solani prin verificarea condițiilor din ultimele 120 de ore. Rezultă o valoare a riscului de la 0 la 100, indicând condiții favorabile pentru boală.
Nu sunt necesare pulverizări atunci când riscul este scăzut. În perioadele cu risc moderat, intervalul de pulverizare poate fi prelungit, iar în perioadele cu risc ridicat, este posibil să trebuiască să fie redus sau să se recomande utilizarea unui compus mai eficient.
În cazul umezelii consecutive a frunzelor, acesta acumulează valori în funcție de temperatură pentru fiecare minut:
- 12 °C până la 15 °C: acumulare de 1 pe minut
- 16 °C până la 17 °C: acumulare de 2 pe minut
- 18°C și mai mult: acumulare de 4 pe minut
La sfârșitul perioadelor de umezeală a frunzelor, se evaluează valorile acumulate:
- Valori > 4096: Riscul crește cu 64 de puncte și se scade 4096 din valoare
- Valori reziduale > 2048: riscul crește cu 16 puncte și se scade 2048 din valoare
- Valori reziduale >1024: Riscul crește cu 4 și scăderea a 1024 din valoare
Dacă radiația globală este consecutiv mai mare de 800 W/m², se acumulează timpul în minute și se evaluează valorile atunci când radiația scade:
- Valoare > 512: Risc - 32 de puncte, valoare - 512
- Valoare > 256: Risc - 8 puncte, valoare - 256
- Valoare > 128: Risc - 2 puncte, valoare - 128
Literatură
- https://cropprotectionnetwork.org/encyclopedia/rhizoctonia-root-rot-of-wheat#:~:text=It%20is%20caused%20by%20Rhizoctonia,result%20in%20premature%20plant%20death.
- https://ahdb.org.uk/knowledge-library/rhizoctonia-stunt-symptoms-and-risk-in-cereals
- https://cropwatch.unl.edu/rhizoctonia-root-rot#:~:text=Disease%20Symptoms&text=These%20reddish%20brown%20lesions%20may,uneven%20because%20of%20stunted%20plants.
- https://extensionaus.com.au/FieldCropDiseasesVic/docs/identification-management-of-field-crop-diseases-in-victoria/soil-borne-diseases/rhizoctonia-root-rot/
- https://www.florimond-desprez.com/es/wp-content/uploads/sites/6/2015/11/rhizoctonia_eng.pdf
Agenți patogeni
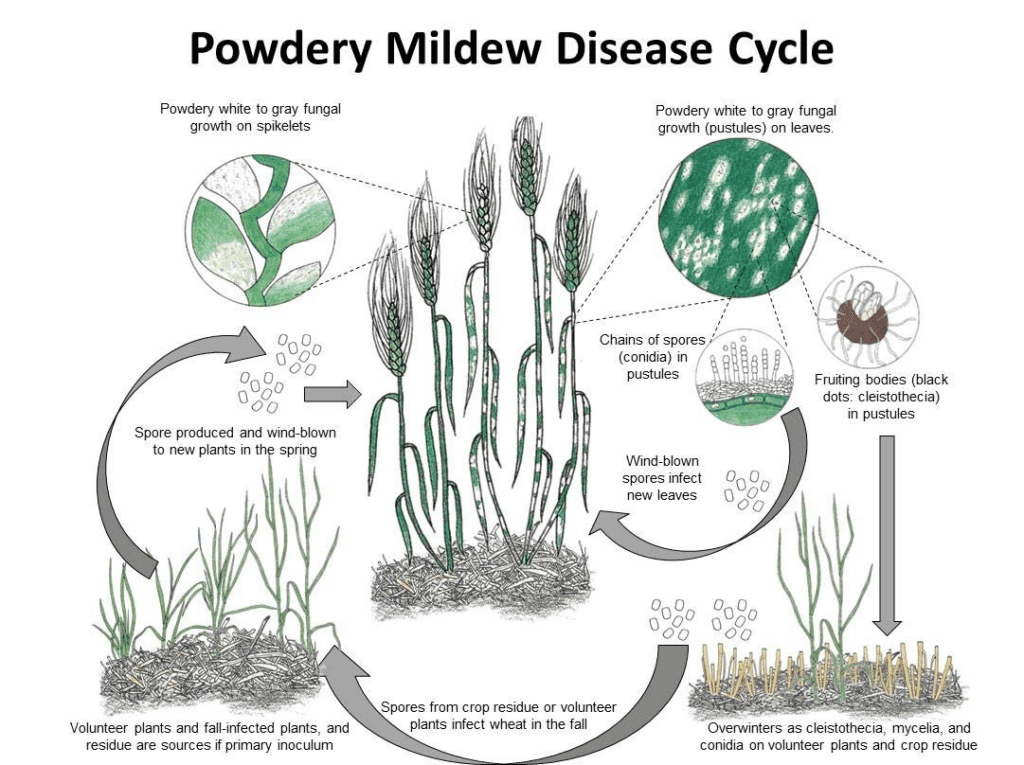
Blumeria graminis este agentul patogen al oidiumului la grâu și orz. Acesta poate ierna sub formă de micelii sau cleistoteci. Primăvara, sporii germinează și infectează plantele gazdă în condiții de răcoare și umiditate, fără a necesita apă liberă pe suprafața plantelor.
Ciuperca se reproduce atât asexuat, cât și sexuat.
În ciclul asexuat, agentul patogen trece prin cicluri rapide de infecție și producție de conidii (spori asexuați), care sunt esențiale pentru răspândirea bolii. Conidiile sunt produse în miceliu la fiecare 7 până la 10 zile în condiții favorabile. Odată dispersate de vânt, aceste conidii aterizează pe o nouă gazdă, germinează și formează appressoria și haustoria pentru a extrage nutrienții din plantă. Conidiile de Blumeria graminis pot germina chiar și la umiditate scăzută și la diferite temperaturi.
În ciclul sexual, ciuperca produce cleistotecii, structuri rezistente care pot supraviețui în condiții adverse. Cleistotecii conțin ascospori (spori sexuali), iar noile infecții sunt inițiate atunci când acești spori sunt eliberați.
Simptomele

Oidioza din grâu și orz afectează producția de boabe prin reducerea numărului de căpițe și diminuarea dimensiunii și greutății boabelor. Cu cât infecția are loc mai devreme, cu atât daunele sunt mai mari.
Orzul prezintă o rezistență legată de vârstă la oidioză. Pe măsură ce orzul se maturizează, în locul petelor tipice de miceliu, pe frunze pot apărea pete negre-maronii bine definite ("pete de smoală").
Grâul este deosebit de sensibil la oidioză între perioada de înfrățire și cea de coacere a laptelui, în special atunci când sunt afectate frunzele pavilionare și cojile. Cu toate acestea, în comparație cu orzul, grâul poate tolera mai bine infecțiile timpurii.
Mucegaiul pulverulent alb-cenușiu se dezvoltă pe frunze, tulpini și capete. Creșterea fungică este mai evidentă pe suprafața superioară a frunzelor, deși se poate dezvolta și pe partea inferioară. Inițial, pustulele par albe, dar devin treptat gri spre maro pe măsură ce se maturizează și se pot coaliza, formând mase mai mari, adesea înconjurate de cloroză. Pe partea opusă a frunzelor infectate, țesuturile devin galbene, devenind ulterior cafenii sau maro. Cleistothecia sunt vizibile, de asemenea, ca puncte mici, rotunde, negre în colonii gri mai vechi. În cazurile severe, frunzele pot muri.
Modelul FieldClimate
Model de mucegai pulverulent al grâului
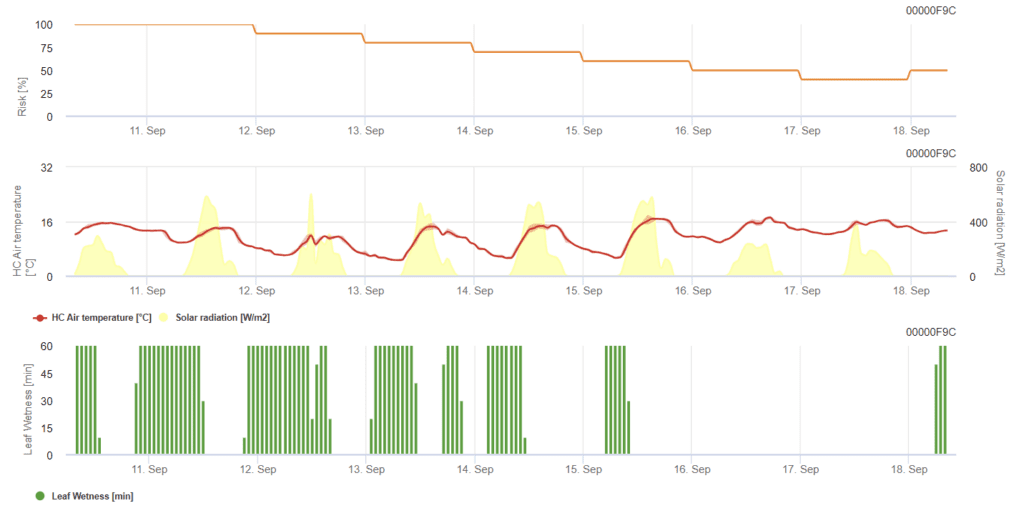
Senzori necesari:
- Temperatura aerului
- Umiditatea frunzelor
- Radiația solară
Modelul determină riscul de apariție a oidiumului, variind pe o scară de la 0 (nicio infecție) la 100% (infecție completă). Modelul este relevant în timpul stadiului BBCH de la 21 la 39 pentru orz și de la 21 la 75 pentru grâu și ovăz.
Stadiul BBCH este o scară de creștere pentru a descrie dezvoltarea plantelor. Există două cifre, scala variază de la 00 la 99 - 00 se referă la tratarea semințelor, iar 99 este momentul în care are loc tratamentul după recoltare. Prima cifră indică stadiul specific de dezvoltare; 0 înseamnă germinare, 1 înseamnă dezvoltarea frunzelor, 2 înseamnă cultivare, 3 înseamnă alungirea tulpinii, 4 înseamnă răsărire, 5 înseamnă apariția inflorescenței și răsărire, 6 înseamnă înflorire și antezis, 7 înseamnă dezvoltarea fructelor, 8 înseamnă coacere și 9 înseamnă senescență.
- BBCH 21: înfrățirea - Începutul înfrățirii; prima înfrățire detectabilă.
- BBCH 39: Alungirea tulpinii - stadiul de frunză steag; frunza steag complet derulată și ligula doar vizibilă.
- BBCH 75: Dezvoltarea fructului - lapte mediu; conținutul de boabe este lăptos și boabele ating dimensiunea finală, dar sunt încă verzi.
Riscul crește în cazul temperaturilor cuprinse între 12 °C și 21 °C în majoritatea orelor din zi și al radiațiilor globale scăzute. Umiditatea frunzelor, radiațiile globale ridicate și temperaturile ridicate de peste 32 °C scad riscul în schimb.
Literatură
- Both, M., & Spanu, P. D. (2004). Blumeria graminis f. sp. hordei, un patogen obligatoriu al orzului. Revista anuală a plantelor, 11, 202-218.
- Cunfer, B. M. (2002). Mucegaiul pulverulent. Grâu pentru panificație: ameliorare și producție, 30, 317-330.
- Der Gräser, E. M. Krankheiten und Schädlinge des Getreides. Lancashire, P. D., Bleiholder, H., Boom, T. V. D., Langelüddeke, P., Stauss, R., Weber, E., & Witzenberger, A. (1991). Un cod zecimal uniform pentru stadiile de creștere ale culturilor și buruienilor. Analele de biologie aplicată, 119(3), 561-601. https://ohioline.osu.edu/factsheet/plpath-cer-11
Agenți patogeni
Există trei rugini și agenți patogeni importanți:
- Frunză/ Rugină brună: Puccinia triticina
- Stem/ Rugină neagră: Puccinia graminis
- Dungă/ Rugină galbenă: Puccinia striiformis
Rugina brună este cea mai frecventă boală a ruginii grâului. Rugina neagră este cunoscută și sub numele de rugina de vară, datorită dezvoltării abundente de teliospori negri strălucitori. Este cea mai devastatoare boală de rugină, cauzând 50 % din pierderi într-o lună în condiții favorabile. Rugina galbenă este o boală a grâului în climatele mai reci, în general asociată cu altitudinile mai mari și latitudinile nordice. Caracteristicile sale sunt urediniosporii de culoare galbenă. În cazuri grave, poate provoca daune la fel de importante ca rugina neagră.
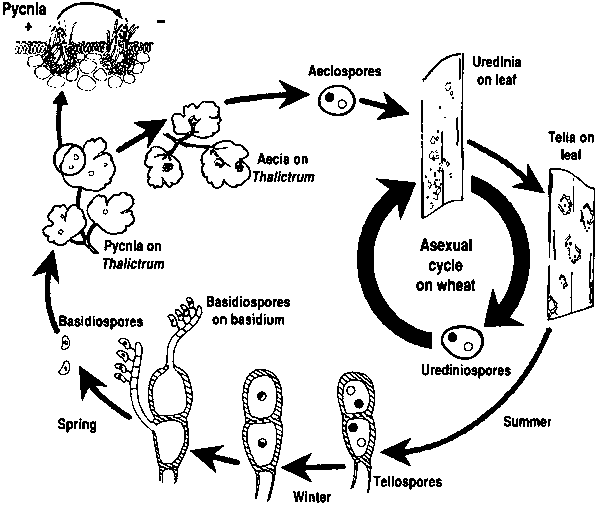
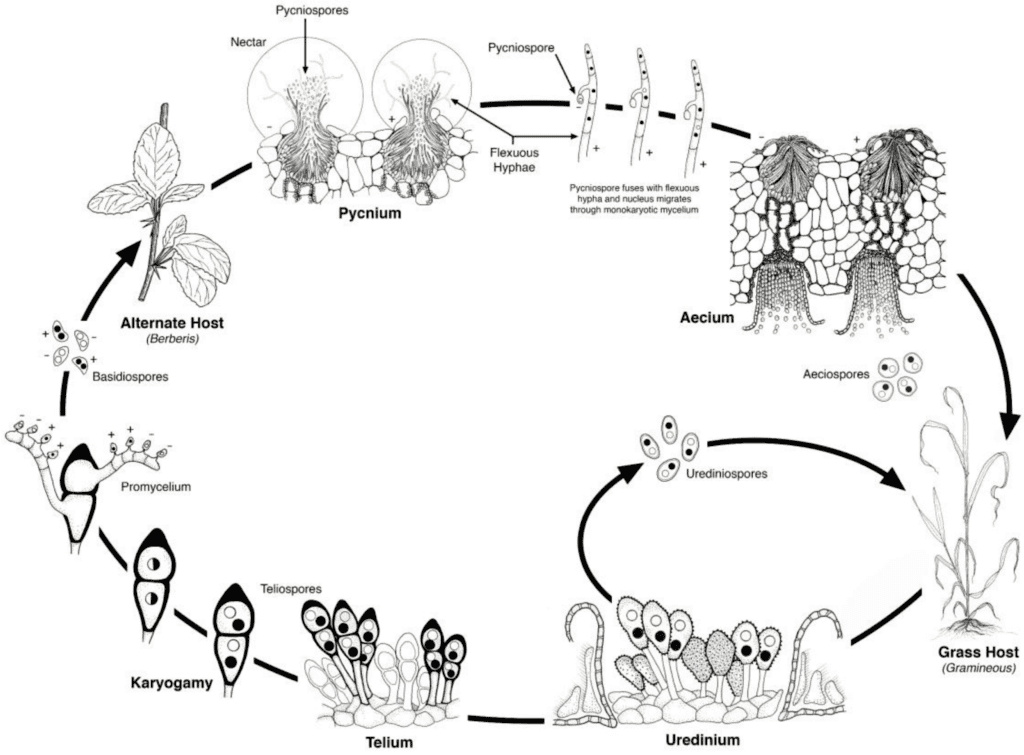
Agenții patogeni ai ruginii grâului au cicluri de reproducere asexuată și sexuală și au nevoie de două plante gazdă - o gazdă economică și o gazdă alternativă. Gazda economică este grâul, iar gazda alternativă este, de obicei, o buruiană sau o plantă nativă, permițând ciupercilor să treacă prin faza sexuală. Puccinia striiformis nu este cunoscută ca având o gazdă alternativă, constând din stadii urediniale și teliale.
În ciclul asexuat, urediniosporii sunt produși pe planta gazdă și pot continua să o infecteze în mod repetat, ducând la o infecție rapidă și generalizată în condiții favorabile.
Ciclul sexual al agentului patogen începe atunci când teliosporii, formați în infecțiile urediniale pe planta gazdă, germinează și sunt supuși meiozei pentru a produce bazidiospori. Aceste bazidiospori sunt dispersate la o gazdă alternativă, unde inițiază infecția, ducând la formarea picniei care conține picniospori și hife receptive. Fertilizarea are loc atunci când pycniosporele de la un tip de împerechere sunt transferate la celălalt prin nectar. În urma fertilizării, pe partea inferioară a frunzelor gazdei alternative se dezvoltă cupe aeciale, care eliberează aeciospori. Acești aeciospori sunt dispersați și infectează noi gazde, după care urmează infecția uredinială, completând ciclul de viață al agentului patogen.
Simptomele

A) Puccinia graminis
B) Puccinia striiformis
C) Puccinia triticina
Puccinia triticina
P. triticina infectează în primul rând lamelele frunzelor, dar poate afecta ocazional tecile frunzelor, glumele și coamele, formând mici pete galbene pe suprafața superioară a frunzelor. Aceste pete se transformă în pustule de culoare portocalie înconjurate de un halou galben pe măsură ce boala progresează. Pustulele produc spori portocalii sau negri, vizibili pe frunze. Spre deosebire de rugina neagră, aceasta produce mai puțini teliospori. Unele plante pot prezenta pete hipersensibile sau uredinia înconjurate de zone clorotice sau necrotice.
Puccinia graminis
P. graminis afectează în principal tulpinile, dar poate infecta și frunzele, tecile, glumele, mugurii și chiar semințele. Inițial, apar leziuni brun-roșcate și, pe măsură ce boala progresează, se formează teliospori negri în interiorul pustulelor. Pe tulpini și pe tecile frunzelor se dezvoltă mai întâi mase de urediniospori roșii-cărămizi și apar mici pete clorotice, care se transformă în pustule liniare sau în formă de diamant, care pot crește în dimensiune. La maturitate, producția de urediniospori se oprește și se produce un strat de teliospori negri, ceea ce face ca tulpinile să apară înnegrite la sfârșitul sezonului. În cazurile grave, infecția slăbește tulpinile plantelor, ceea ce duce la cădere.
Puccinia striiformis
Trăsătura caracteristică a P. striiformis este prezența de pustule galbene cu laturi drepte. TJey apar în dungi înguste, alungite, de diferite lungimi, adesea însoțite de cloroză și necroză. Pe măsură ce boala se maturizează, sporii galben-portocalii se dezvoltă în interiorul pustulelor, iar țesuturile din jur devin maro și se usucă, creând un aspect ars.
Modelul FieldClimate
Modelul ruginei Puccinia
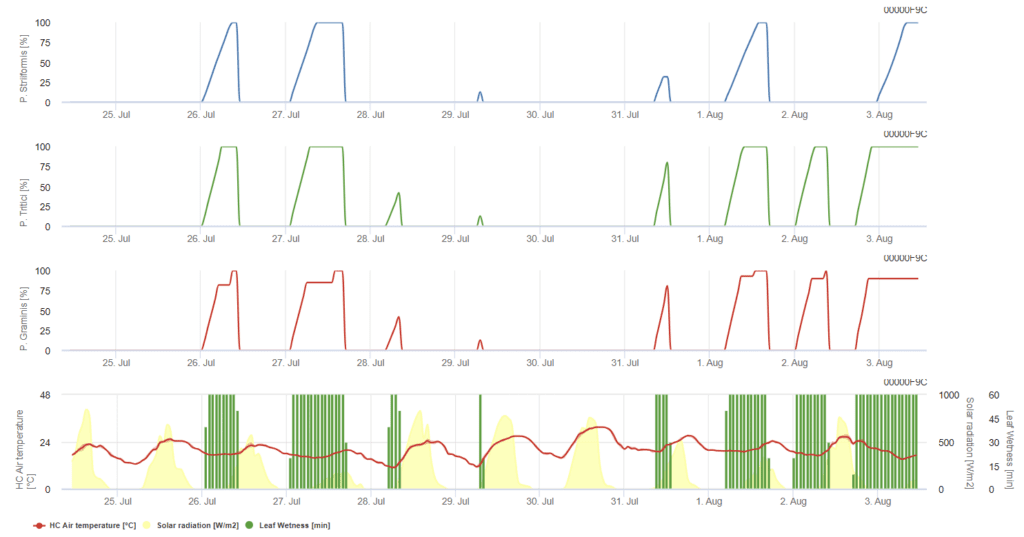
P. triticina, P. graminis, P. striiformis sunt prezentate fiecare în trei grafice colorate diferite.
Model de infecție cu Puccinia triticna
Modelul presupune că infecția necesită o temperatură orară acumulată de 90°C de umezeală a frunzelor într-un interval de temperatură a aerului de la 5°C la 30°C. Condițiile sunt similare cu P. graminis, dar cu un prag de temperatură mai scăzut de 5°C.
Dacă se constată o infecție la 100%, trebuie luată în considerare o măsură fitosanitară curativă. În cazul în care riscul este de 80%, iar prognoza meteo prevede mai multe perioade de umezeală a frunzelor, ar putea fi luate măsuri de protecție a frunzelor.
Umiditatea frunzelor pentru temperaturile medii orare acumulate pentru 90°C:
- (dacă T <= 22,5°C atunci ∑(Th) altfel ∑ (22,5-(Th-22,5))
- 5°C < Temp. < 30°C
Model de infecție cu Puccinia graminis
Infecțiile au loc după câteva ore de umezeală a frunzelor în condiții optime de temperatură. Modelul presupune că infecția are nevoie de o temperatură orară acumulată de 80°C de umezeală a frunzelor într-un interval de temperatură a aerului de la 10°C la 35°C. Ea preferă o temperatură puțin mai ridicată decât P. triticina iar infecția trebuie să fie urmată de lumina soarelui.
Atunci când se constată o infecție cu 100%, condițiile au fost favorabile pentru ciupercă și, prin urmare, ar trebui luate în considerare măsurile de protecție a plantelor.
Umiditatea frunzelor pentru temperaturi medii orare cumulate de 80°C, urmată de o perioadă de lumină (150 W/m²) pentru temperaturi medii orare cumulate de 30°C:
- (if T <= 24°C then ∑(Th) else ∑ (Th-24))
- 10°C < Temp. < 35°C
Model de infecție cu Puccinia striiformis
Puccinia striiformis este rugina grâului din zonele cu climă răcoroasă, temperatura sa optimă fiind deja de 15°C. Infecțiile au loc după câteva ore de umezeală a frunzelor în condiții de temperatură optimă. Modelul presupune că infecția necesită o temperatură orară acumulată de 80°C de umezeală a frunzelor într-un interval de temperatură a aerului de la 5°C la 20°C. Nu există infecții în perioadele cu intensitate luminoasă scăzută.
Înainte ca graficul să ajungă la 100%, ar putea fi efectuată o aplicare de protecție, în timp ce după aceea ar trebui luate în considerare strategii de protecție curative (sistemice).
Umiditatea frunzelor și lumina pentru temperaturile medii orare acumulate pentru 80°C:
- (if T <= 15°C then ∑(Th) else ∑ (Th-15))
- 5°C < Temp. < 20°C
Literatură
- Bolton, M. D., Kolmer, J. A. și Garvin, D. F. (2008). Rugina frunzelor de grâu cauzată de Puccinia triticina. Patologia moleculară a plantelor, 9(5), 563-575.
- Chen, X. M. (2005). Epidemiologia și controlul ruginii striate [Puccinia striiformis f. sp. tritici] la grâu. Revista canadiană de patologie vegetală, 27(3), 314-337.
- Figueroa, M., Hammond-Kosack, K. E., & Solomon, P. S. (2018). O revizuire a bolilor grâului - o perspectivă de teren. Patologia moleculară a plantelor, 19(6), 1523-1536.
- Guide, A. Bolile ruginii frunzelor, tulpinilor și dungilor la grâu. Leonard, K. J., & Szabo, L. J. (2005). Rugina tulpinilor de cereale mici și graminee cauzată de Puccinia graminis. Patologia moleculară a plantelor, 6(2), 99-111.
- Kolmer, J. (2013). Rugina frunzelor de grâu: biologia agenților patogeni, variația și rezistența la gazdă. Păduri, 4(1), 70-84.
- Singh, R. P., Huerta-Espino, J., Roelfs, A. P., & Curtis, B. C. (2002). Rugina grâului. Creștere, 2(25), 35.