
Verificați ce set de senzori este necesar pentru monitorizarea bolilor potențiale ale acestei culturi.

Agenți patogeni
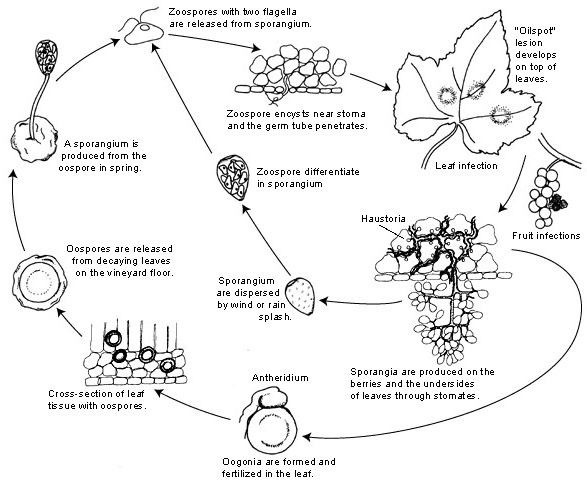
Agentul patogen al mucegaiului pufos al strugurilor este Plasmopara viticola. Este un parazit obligatoriu, ceea ce înseamnă că are nevoie de organe de viță de vie verzi și proaspete pentru creștere. Are atât stadii de reproducere asexuată, cât și sexuală -zoospori și oospori. Prin urmare, există două etape de infecție: infecția primară și infecția secundară.
Infecția primară începe prin germinarea oosporilor care iernează, formând sporangele primăvara, când vremea devine caldă. Sporangele sunt produse noaptea, deoarece lumina soarelui inhibă sporularea. Oosporii cu pereți groși ajută agentul patogen să supraviețuiască condițiilor grele de iarnă și să inițieze infecția primară. Sporangele sunt dispersate de ploaie sau de vânt și eliberează zoospori în apa liberă. Pentru ca infecția să aibă succes, este necesară umezirea frunzelor sau precipitații suficient de lungi.
Infecția secundară este posibilă numai în prezența petelor uleioase mature și se produce prin zoospori și sporange. Zoosporii și sporangele sunt foarte sensibile la umiditatea scăzută și la lumină, ceea ce le scurtează viabilitatea, ceea ce înseamnă că cea mai mare parte a infecției are loc imediat după eliberare. Zoosporii pătrund în țesuturile plantelor prin intermediul tuburilor germinale și dezvoltă hife pentru continuarea infecției. Aceste hife formează leziuni uleioase după perioada de incubație care variază între 5 și 21 de zile în funcție de temperatură - cu cât temperatura este mai scăzută, cu atât durează mai mult.
Pe măsură ce vremea se răcește din nou, ceea ce este nefavorabil pentru patogen, acesta trece la producția sexuală, producând oospori. Acești oospori rămân latenți în timpul iernii, iar ciclul se repetă.
Simptomele

Pe suprafața superioară a frunzelor apar mici leziuni galbene, uneori înconjurate de halouri maronii. Petele de ulei se extind și se integrează pe măsură ce maturarea și halourile dispar. Sporangele produc ulterior un mucegai alb-cottoniu pe partea inferioară a leziunilor și urmează necroza. Zone maro uleioase apar și pe lăstari, tulpini și fructe de pădure. În timpul nopților calde și umede, acestea se pot acoperi cu puf alb din cauza sporulării. Infecția severă provoacă căderea prematură.
Modele FieldClimate
- Există două modele în FieldClimate - infecție primară și secundară.
- Primul grafic prezintă timpul de incubație după finalizarea unei infecții. Graficele inferioare descriu evoluția infecției pentru un nivel de gravitate slab, moderat și grav. Se presupune că există o infecție atunci când curba crescătoare din grafic ajunge la 100% - pulverizarea poate fi luată în considerare dacă se indică un nivel grav de infecție.
Infecție primară
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umiditatea frunzelor
- Precipitații
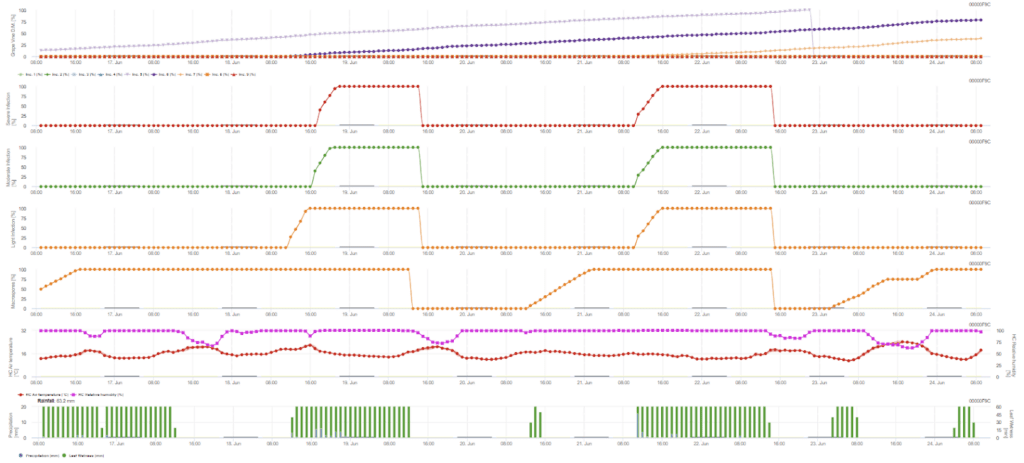
Infecția primară verifică dacă vremea este potrivită pentru dezvoltarea sporangiilor. Acesta este cazul atâta timp cât frunzele sunt umede sau umiditatea relativă după ploaie nu scade sub 70%. Sporangele se pot dezvolta în decurs de 16 până la 24 de ore, în funcție de temperatură. O ploaie continuă de 5 mm este interpretată ca o ploaie puternică care poate răspândi zoosporii.
Infecție secundară
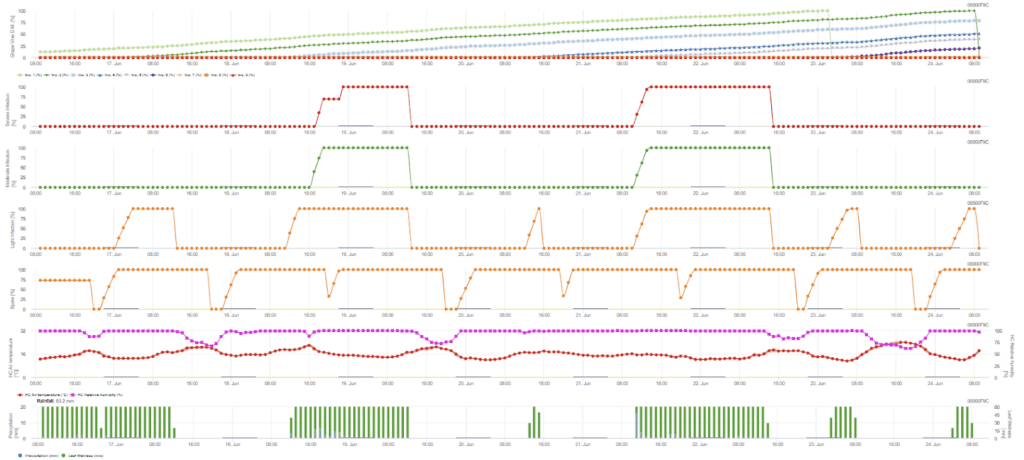
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umiditatea frunzelor
- Precipitații
Infecția secundară se verifică dacă temperatura este mai mare de 12 °C și umiditatea relativă este mai mare de 95%. Rata de producere a sporangiilor crește cu temperaturi de până la 23 °C. Dacă această condiție durează pentru o temperatură orară cumulată de peste 50 °C, presupunem că sporulația este terminată și că există sporange noi în podgorie. Cele 50 °C acumulate corespund, de exemplu, la 4 ore cu 13 °C sau 3 ore cu 17 °C. Sporangele mor rapid atunci când este mai cald și mai uscat - atunci când umiditatea relativă scade sub 50%, o resetăm la 0, iar atunci când temperatura depășește 29 °C, nu poate avea loc nicio sporulație.
Literatură
- Ash, G. (2000). Downy mildew al strugurilor. The Plant Health Instructor.
- Gessler, C., Pertot, I. și Perazzolli, M. (2011). Plasmopara viticola: o trecere în revistă a cunoștințelor privind pucioasa viței de vie și gestionarea eficientă a bolii. Phytopathologia Mediterranea, 50(1), 3-44.
- Kennelly, M. M., Gadoury, D. M., Wilcox, W. F., Magarey, P. A. și Seem, R. C. (2007). Infecția primară, productivitatea leziunilor și supraviețuirea sporangiei la agentul patogen Plasmopara viticola al mucegaiului viței de vie. Phytopathology, 97(4), 512-522.
- Koledenkova, K., Esmaeel, Q., Jacquard, C., Nowak, J., Clément, C. și Ait Barka, E. (2022). Plasmopara viticola, agentul cauzal al pucioasei viței de vie: de la taxonomia sa la gestionarea bolii. Frontiers in Microbiology, 13, 889472.
Agenți patogeni
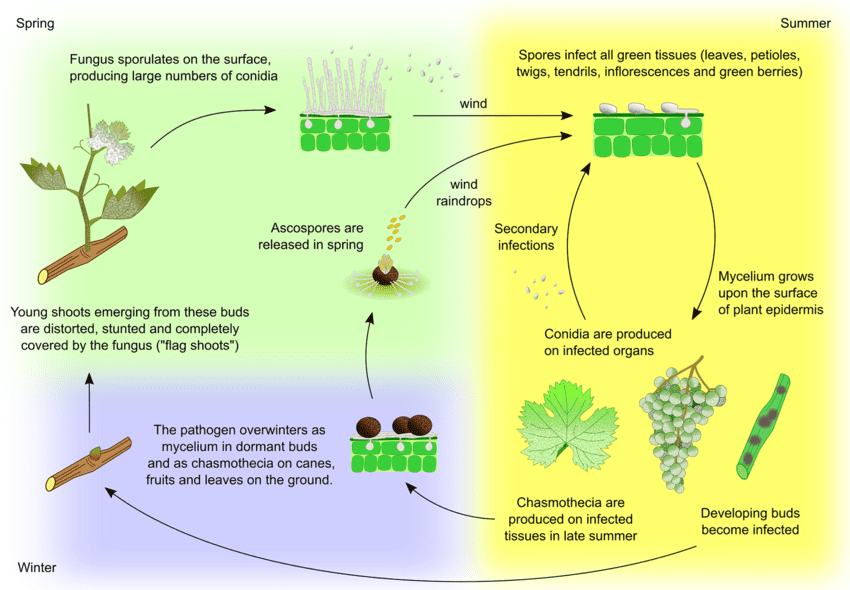
Agentul patogen al oidiumului la struguri este Uncinula necator. Există două surse principale de inocul - miceliul care iernează și ascosporii din cleistoteci - a căror importanță poate varia în funcție de regiune.
Miceliul cauzează lăstari pavilionari în primăvară. Lăstarii steag sunt lăstari parțial sau complet acoperiți de mucegai, formați din muguri infectați latenți. Pe măsură ce se formează colonii în interior, acești lăstari sunt bine vizibili și au adesea frunze deformate.
Cleistothecia eliberează ascospori la începutul primăverii. Precipitațiile joacă un rol important în eliberarea ascosporelor. Pe măsură ce temperatura crește, dehiscența are loc mai frecvent. Ascosporii germinează în termen de 12 ore, formând tuburi germinative și appressorium pe țesuturile plantelor, ducând la formarea de colonii dispersate.
Infecția secundară are loc prin conidii produse în colonii. La fel ca ascosporii, acestea germinează și dezvoltă tuburi germinative și appressorium. U. necator nu are nevoie de apă liberă pentru infectare și nici de umiditate relativă ridicată pentru formarea conidiei. Atunci când condițiile meteorologice devin mai puțin favorabile, ciuperca formează cleistoteci. Aceasta iernează fie supraviețuind în cleistoteci, fie sub formă de miceliu vegetativ în mugurii infectați inactivi, iar ciclul se repetă.
Simptomele

Simptomele pot fi găsite pe toate părțile verzi ale viței de vie. Lăstarii dezvoltă colonii și se acoperă cu miceliu alb-griuriu, cunoscut sub numele de "lăstari steag". Acestea produc frunze deformate. Coloniile de mucegai se găsesc și pe frunze - fie pe partea inferioară, fie pe ambele părți. Boabele prezintă pete cenușii și se despică în cazurile grave. Tulpinile prezintă pete de culoare roșu-brun închis și pot muri de la vârfuri.
Modele FieldClimate
Infecția cu ascospori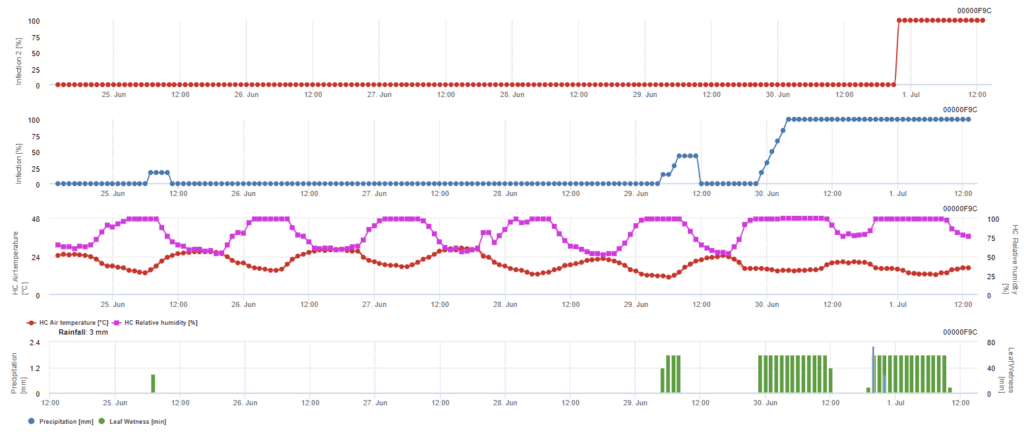
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umezeala frunzelor
- Precipitații
Acest model este recomandat atunci când ascosporii se formează în timpul iernii reci (deci nu sunt infectați de miceliu). Infecția cu ascospori prezice eliberarea de ascospori și infecția inițială pe baza temperaturilor medii din timpul perioadelor prelungite de umezeală a frunzelor. Pentru calcul, sunt necesari aproximativ 2,5 mm de precipitații pentru eliberarea ascosporilor, urmate de un minim de 8-12 ore de umezeală a frunzelor și temperaturi cuprinse între 10 și 15°C. Odată ce infecția a avut loc, modelul trece la o fază de evaluare a riscului de boală (modelul californian de risc) și se bazează pe efectele temperaturii asupra ratei de reproducere a agentului patogen.
Modelul de risc californian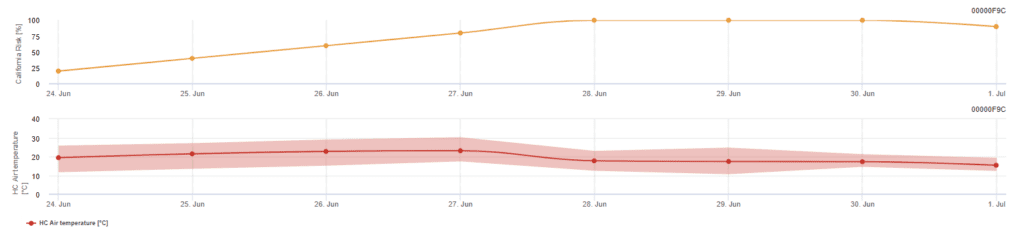
Senzori necesari:
- Temperatura aerului
Modelul se bazează pe studii de laborator efectuate în California. După eliberarea și germinarea ascosporelor (model), dezvoltarea și reproducerea ulterioară a oidiozei sunt influențate de temperaturi. Modelul evaluează temperaturile și apreciază riscul de dezvoltare a ocidului prin utilizarea unui indice de la 0 la 100 de puncte.
Trei zile la rând cu minimum șase ore consecutive de temperatură între 21 și 30'c sunt necesare pentru a iniția indicele de evaluare a riscurilor. Se câștigă 20 de puncte pentru fiecare zi care îndeplinește 6 sau mai multe ore consecutive între 21 și 32°C și se pierd 10 puncte pentru cele care nu fac acest lucru sau atunci când temperatura depășește 32°C sau coboară sub 21°C.
Valorile scăzute ale indicelui de 0~30 indică faptul că agentul patogen nu se reproduce. Un indice de 40~50 este considerat moderat și implică o rată de reproducere a oidului praf de aproximativ 15 zile. Valorile indicelui de peste 60 indică faptul că agentul patogen se reproduce rapid (la fiecare 5 zile) și că riscul unei epidemii este mare.
Modelul de risc Pessl Instruments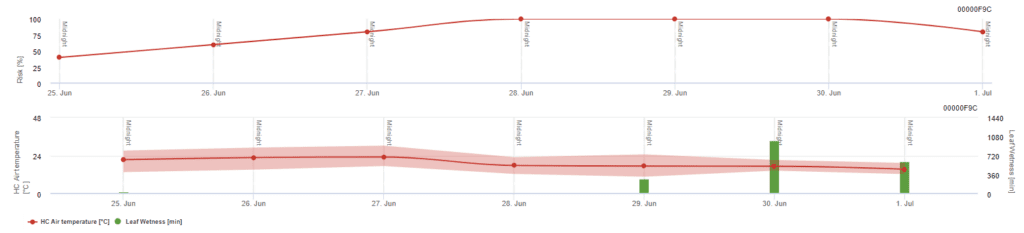
Senzori necesari:
- Temperatura aerului
- Umiditatea frunzelor
Pe lângă temperatura aerului, umezeala frunzelor poate fi un factor de orientare, deoarece perioadele lungi de umezeală a frunzelor conduc la instalarea unei ciuperci antagoniste (Ampelomyces quisqualis), care duce la o scădere a U. necator. Principiile de bază ale modelului sunt echivalente cu cele ale modelului californian de risc, însă umezeala foliară este, de asemenea, luată în considerare în acest model. Umiditatea frunzelor mai mare de 8 ore duce la o scădere de 10 puncte.
Valorile scăzute ale indicelui de 0~20 indică faptul că agentul patogen nu se reproduce. Un indice de 20~60 este considerat moderat și un interval normal de pulverizare este valabil. Valorile indicelui de peste 60 indică faptul că agentul patogen se reproduce rapid (la fiecare 5 zile) și că riscul unei epidemii este mare, astfel încât se recomandă scurtarea intervalului de pulverizare.
Literatură
- Gadoury, D. M., & Pearson, R. C. (1990). Dehiscența ascocarpului și evacuarea ascosporilor la Uncinula necator. Phytopathology, 80(4), 393-401.
- Gadoury, D. M., & Pearson, R. C. (1990). Germinarea ascosporilor și infecția lui Vitis de către Uncinula necator. Phytopathology, 80(11), 1198-1203.
- Hall, T. W. (2000). Epidemiologia mucegaiului praf al strugurilor, Uncinula necator, în Willamette Valley.
- Halleen, F., & Holz, G. (2001). O privire de ansamblu asupra biologiei, epidemiologiei și controlului Uncinula Necator (mucegaiul pulverulent) pe vița de vie, cu referire la Africa de Sud. South african journal of Enology and Viticulture, 22(2), 111-121.
- Rügner, A., Rumbolz, J., Huber, B., Bleyer, G., Gisi, U., Kassemeyer, H. H., & Guggenheim, R. (2002). Formarea structurilor de iernare ale Uncinula necator și colonizarea viței de vie în condiții de câmp. Patologia plantelor, 51(3), 322-330.
Agenți patogeni
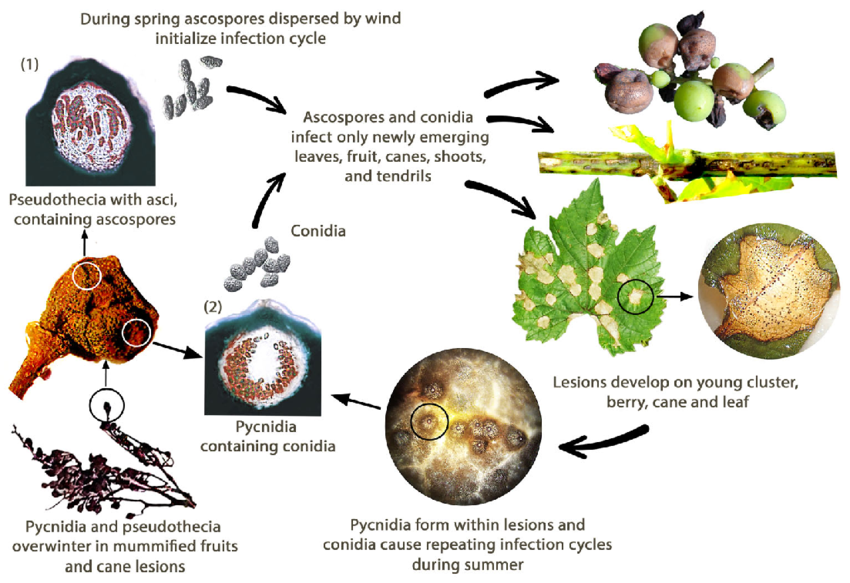
Putregaiul negru al viței de vie este cauzat de ciuperca Guignardia bidwellii. Ciuperca iernează în diferite părți ale viței de vie și poate supraviețui timp de peste doi ani în leziunile lăstarilor infectați.
Primăvara, picnidia produce conidii (spori asexuați), iar pseudotecia generează ascospori (spori sexuali). Conidiile se dispersează de obicei pe distanțe scurte, în timp ce ascosporii pot infecta pe distanțe mai mari. Acești spori sunt transportați de vânt și ploaie pentru a infecta țesuturile tinere ale gazdelor sensibile, inițiind infecții primare. Conidiile servesc și ca inocul secundar, contribuind la răspândirea rapidă și repetată a bolii.
În cursul lunii august, picnidiile se transformă într-un stadiu de iernare care produce pseudotecii, care generează ascospori. Acești ascospori sunt surse importante pentru infecțiile primare din primăvara următoare.
Simptomele

Se dezvoltă pete brun-roșcate care în cele din urmă fuzionează, adesea înconjurate de mici puncte negre, care sunt structuri fungice cunoscute sub numele de picnidii care produc conidii. Frunzele tinere și cu creștere rapidă sunt deosebit de sensibile la această infecție.
Infecțiile fructelor apar de obicei după căderea caliciului, cele mai multe simptome manifestându-se atunci când fructul este la jumătatea sau aproape de dimensiunea maximă. Inițial, apar pete mici, înconjurate de un inel maro, care apoi se măresc și se întunecă, acoperind în cele din urmă întreaga boabă pe măsură ce boala se dezvoltă. În câteva zile, boabele infectate pot deveni mumificate, se pot sparge sau pot cădea prematur.
În plus, sunt afectate și alte părți ale plantei, cum ar fi lăstarii, tulpinile și vreascurile. Se dezvoltă leziuni de formă ovală de culoare purpurie până la neagră, iar picnidiile se împrăștie pe suprafața acestor leziuni.
Model FieldClimate
Viticultură Model de putregai negru
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umiditatea frunzelor
- Precipitații
Modelul se bazează inițial pe literatura publicată de Spotts, dar cu modificarea lui Daniel Molitor. Revizuirea a introdus trei clase de severitate (ușoară, moderată și severă), asemănătoare cu modelele de scabie a mărului. La temperaturi ridicate, perioada de umezeală a frunzelor de 8 ore este suficientă pentru a favoriza infecțiile și se presupune că o infecție este finalizată atunci când un grafic atinge 100%.
Gravitatea infecției depinde de durata de timp a perioadei de temperatură optimă și de umezeală a ciupercii. Infecțiile care îndeplinesc criteriile Spotts sunt considerate infecții ușoare. Infecțiile care îndeplinesc criteriile Spott prin 150% și 200% sunt considerate infecții moderate și, respectiv, severe.
În funcție de presiunea infecției, infecțiile trebuie acoperite preventiv sau trebuie aplicat un spray curativ la scurt timp după infecție. În climatul semiarid moderat din Mosel sau din majoritatea zonelor viticole austriece, pulverizarea după depistarea unei infecții ușoare nu ar fi recomandată.
Literatură
- Wilcox, Wayne F. "Putregaiul negru Guignardia bidwellii". Fișa de identificare a bolii nr. 102GFSG-D4. 2003. Cornell. 24 oct. 2010
- http://www.nysipm.cornell.edu/factsheets/grapes/diseases/grape_br.pdf
- Ellis, Michael A. "Fact sheet Agricultural and Natural Resources: Putregaiul negru al strugurilor". Departamentul de patologie vegetală. The University of Ohio State Extension. 2008
- http://ohioline.osu.edu/hyg-fact/3000/pdf/HYG_3004_08.pdf
- Molitor, D. (2009). Untersuchungen zur Biologie und Bekämpfung der Schwarzfäule (Guignardia bidwellii) an Weinreben. Gesellschaft zur Förderung der Forschungsanstalt Geisenheim.
- Ries, S. M. (1999). Rapoarte privind bolile plantelor: Putregaiul negru al strugurilor. Integrated Pest Management at the Uni. of Illinois. http://ipm. illinois. edu/diseases/series700/rpd703.
- Spotts, R. A. (1977). Efectul duratei umezelii frunzelor și al temperaturii asupra infectivității Guignardia bidwellii pe frunzele de struguri. Phytopathology, 67(11), 1378-1381.
- Szabó, M., Csikász-Krizsics, A., Dula, T., Farkas, E., Roznik, D., Kozma, P., & Deák, T. (2023). Putregaiul negru al strugurilor (Guignardia bidwellii)-o privire de ansamblu. Horticulturae, 9(2), 130.
- http://extension.cropsciences.illinois.edu/fruitveg/pdfs/771-BlackRotOfGrape.pdf
- https://www.missouribotanicalgarden.org/gardens-gardening/your-garden/help-for-the-home-gardener/advice-tips-resources/insects-pests-and-problems/diseases/fruit-spots/black-rot-of-grapes
Agenți patogeni

Lobesia botrana În funcție de regiune, molia are de obicei două până la trei generații pe an. Prima generație (mai și iunie) afectează numai florile, astfel încât tratamentul este necesar numai dacă populațiile de molii sunt foarte mari. A doua generație (iulie și august) și a treia generație provoacă cele mai mari daune, a treia generație fiind deosebit de dăunătoare deoarece coincide cu coacerea strugurilor. O a patra generație poate apărea în climatele mai calde, dar tratamentul nu este, în general, necesar, deoarece coincide cu recoltarea strugurilor.
Molia iernează ca pupă în coconi, iar când temperaturile cresc primăvara, apar adulții din prima generație, masculii apărând de obicei înaintea femelelor. Zborul primei generații începe aproape de răsărirea mugurilor și durează 4-5 săptămâni, timp în care are loc împerecherea. După una sau două zile de la împerechere, femelele depun între 80 și 160 de ouă.
Ouăle, cu un diametru de 0,6 până la 0,8 mm, sunt vizibile cu ochiul liber. Inițial alb-crem, ele devin galbene pe măsură ce embrionul se dezvoltă, cu culoarea neagră a capului larvei. Ouăle eclozează după 66 de grade-zi Celsius (DDC), aproximativ între 7 și 11 zile. Larvele țes părțile florii împreună, dezvoltarea durând între 20 și 30 de zile. Urmează puparea, iar adulții ies la suprafață după 6-14 zile.
"Gradul-zi" se calculează prin măsurarea cu cât temperatura medie zilnică depășește un anumit prag de temperatură de bază necesar pentru dezvoltare. Pe măsură ce temperaturile cresc peste nivelul de bază, gradele-zile se acumulează.
Moliile adulte au o lungime de aproximativ 6-8 mm și o anvergură a aripilor de 11-13 mm. Femelele sunt puțin mai mari decât masculii, deși ambele au aripi similare, cu model mozaicat.
Ouăle din a doua și a treia generație eclozează mai repede, în 3-5 zile. Femelele depun ouăle individual pe fructele umbrite, iar la eclozare, larvele intră în fructe și le scobesc. Toamna, nopțile mai lungi de 11 ore declanșează diapauza, un stadiu de repaus care permite pupeilor să tolereze mai bine frigul, permițându-le să ierneze.
Simptomele
Omizile din prima generație se hrănesc cu inflorescențele viței de vie, scobind bobocii florali și rotindu-i pentru a-i proteja, ceea ce duce la deteriorarea ciorchinilor. În a doua generație, larvele atacă boabele în dezvoltare, creând puncte de intrare pentru agenți patogeni fungici precum Botrytis cinerea. Această hrănire directă provoacă daune vizibile, marcate de obicei de pete întunecate în jurul locurilor de hrănire. Cea de-a treia generație provoacă daunele cele mai semnificative, deoarece larvele pătrund și se hrănesc cu fructe coapte.
Rămân boabele zbârcite și excrementele larvelor. Resturi de țesut epidermic al boabelor rămân lipite de pedicel, împreună cu "pielea" boabelor uscată și scobită.
Model FieldClimate
Modelul moliei strugurilor de viță de vie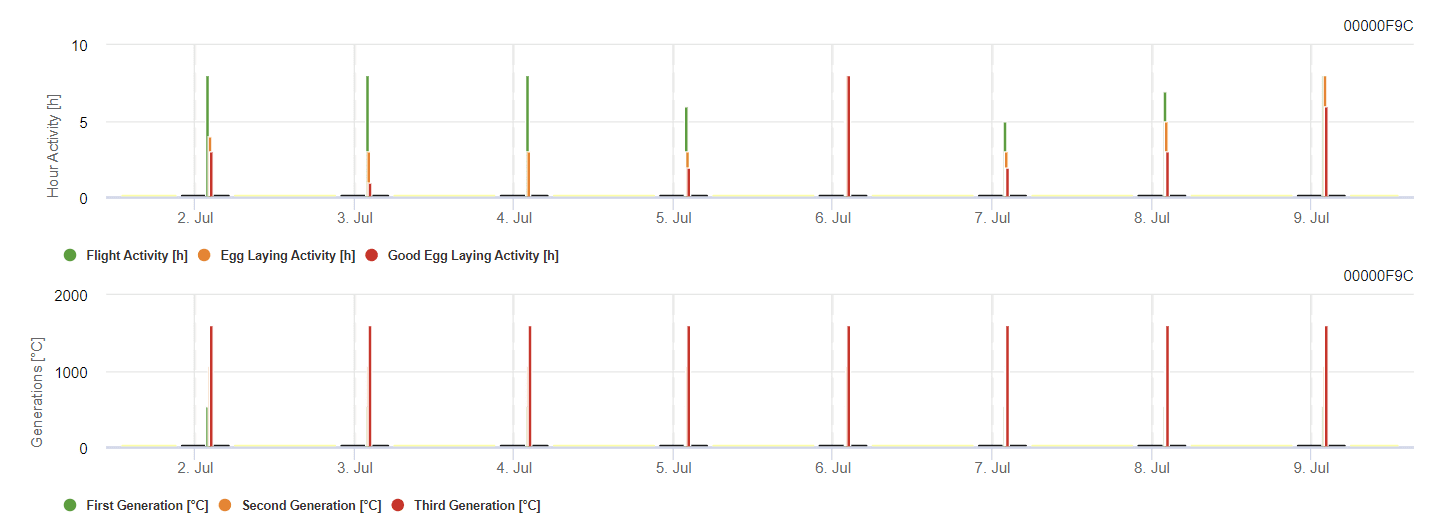
Senzor necesar:
- Temperatura aerului
Modelul calculează riscul în funcție de temperatura aerului. Primul grafic arată perioadele de activitate de zbor cu depunere a ouălor și activitățile bune de depunere a ouălor pentru insectă. Al doilea grafic arată la ce generație trebuie să ne așteptăm în perioada respectivă. În al treilea grafic, sunt afișate datele climatice, care arată acumularea de grade-zile (temperaturi > 8°C până la 24°C pe oră împărțite la 24).
Literatură
- Varela, L. G., Smith, R. J., Cooper, M. L., & Hoenisch, R. W. (2010). Molia europeană a viței de vie, Lobesia botrana. Napa Valley vineyards. Pract. Winery Vineyard, 2010, 1-5.
- https://ipm.ucanr.edu/invasive-and-exotic-pests/european-grapevine-moth/
- https://www.ages.at/en/plant/plant-health/pests-from-a-to-z/grape-berry-moth
Agenți patogeni
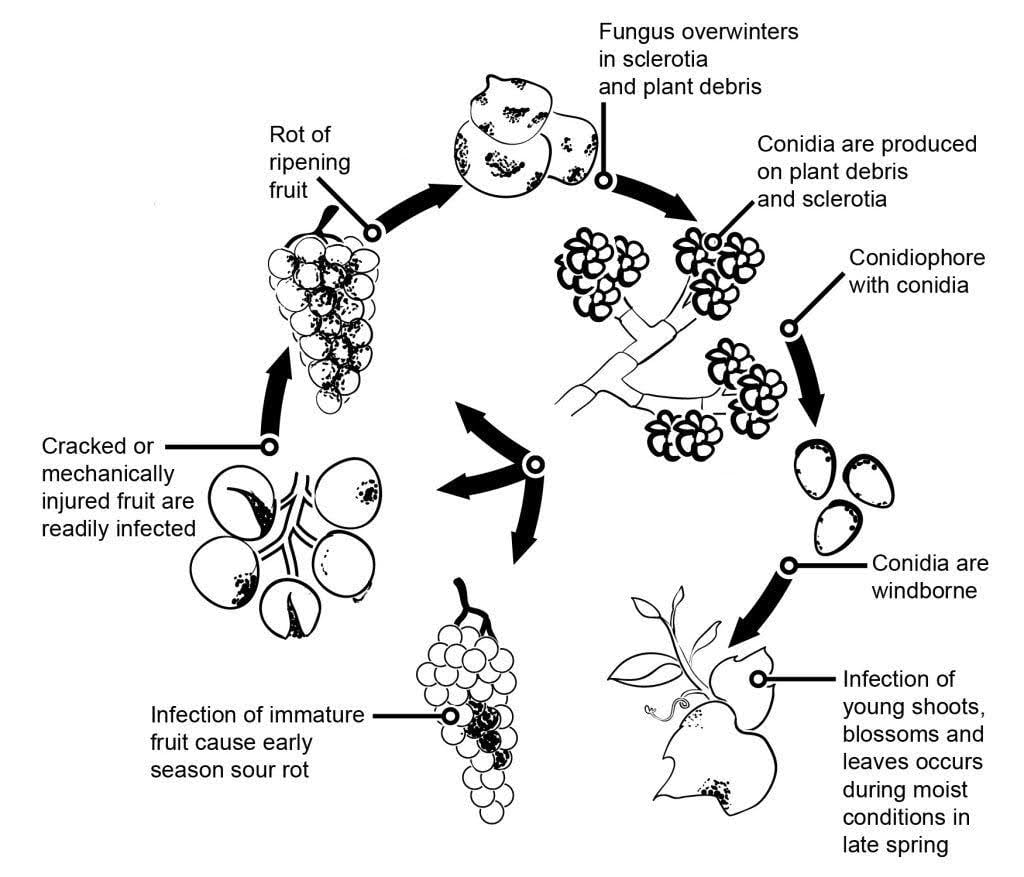
Agentul patogen al mucegaiului cenușiu din vița de vie este Botrytis cinerea.
Epidemiologia mucegaiului cenușiu este influențată în mod semnificativ de infecțiile latente inițiate de infecțiile florilor. Au fost identificate diferite căi de transmitere de la floare la fruct: Ciuperca infectează stiloul florii și ajunge la ovul, unde rămâne latentă datorită mecanismului de apărare preformat al plantei. Infecțiile prin stamine, petale și sepale sunt, de asemenea, esențiale. B. cinerea poate infecta staminele și crește sistematic spre receptacul, răspândindu-se la pedicel și la țesuturile vasculare din boabe. Alte căi implică, de asemenea, creșterea saprofitică a agentului patogen.
Miceliile sau scleroții care iernează sunt principalele surse de infecție în primăvară. Conidiile (sporii asexuați) sunt produse și dispersate de vânt și ploaie. La aterizare, conidiile germinează, formând tuburi germinative și appresorii care penetrează și infectează planta. Ciuperca poate rămâne latentă până când fructul se coace și conținutul de zahăr crește. Conidiile generate de sursele de inocul primar urmează un ciclu diurn de inițiere, producție și diseminare.
Deși stadiul sexual al B. cinerea este rar observat, agentul patogen devine mai activ pe măsură ce boabele se coc. Conținutul crescut de zahăr din fructe crește susceptibilitatea la infecție. În același mod, florile sunt foarte sensibile atunci când se ofilesc, polenul abundent crescând severitatea infecției.
Simptomele

Mucegaiul gri infectează în primul rând boabele de struguri coapte, care inițial par moi și îmbibate cu apă. În timp, boabele devin brun-roșcate și se zbârcesc. În condiții favorabile, acestea se acoperă cu mase gri de micelii fungice și conidii. Boabele sănătoase pot fi infectate și prin contact direct cu cele bolnave.
Ciuperca poate provoca, de asemenea, o arsură a florilor, care poate duce la pierderi semnificative de recoltă la începutul sezonului. Deși infecțiile foliare sunt rare, atunci când apar, acestea încep ca pete verzi terne de-a lungul nervurilor, care în cele din urmă se transformă în leziuni necrotice.
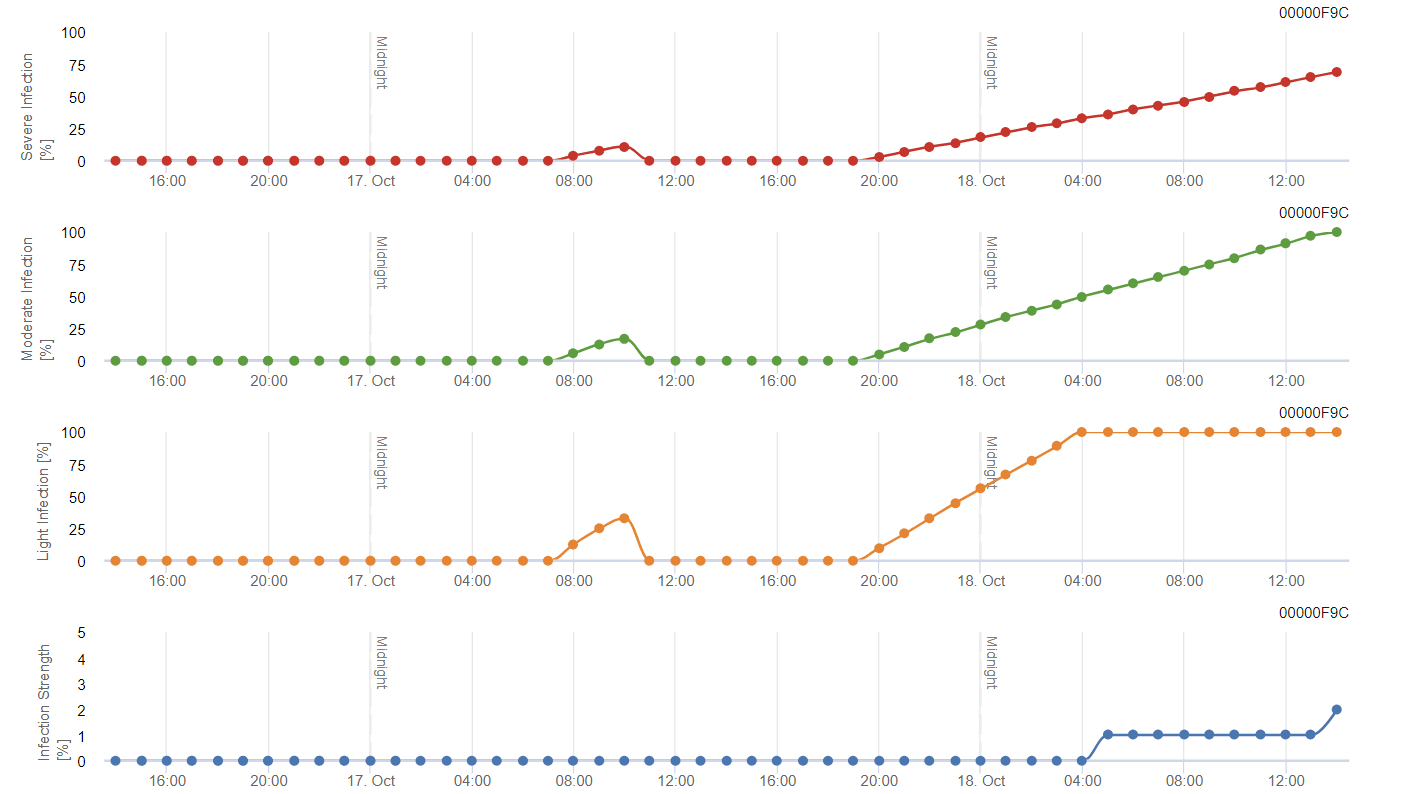
Model FieldClimate
Model general de matriță gri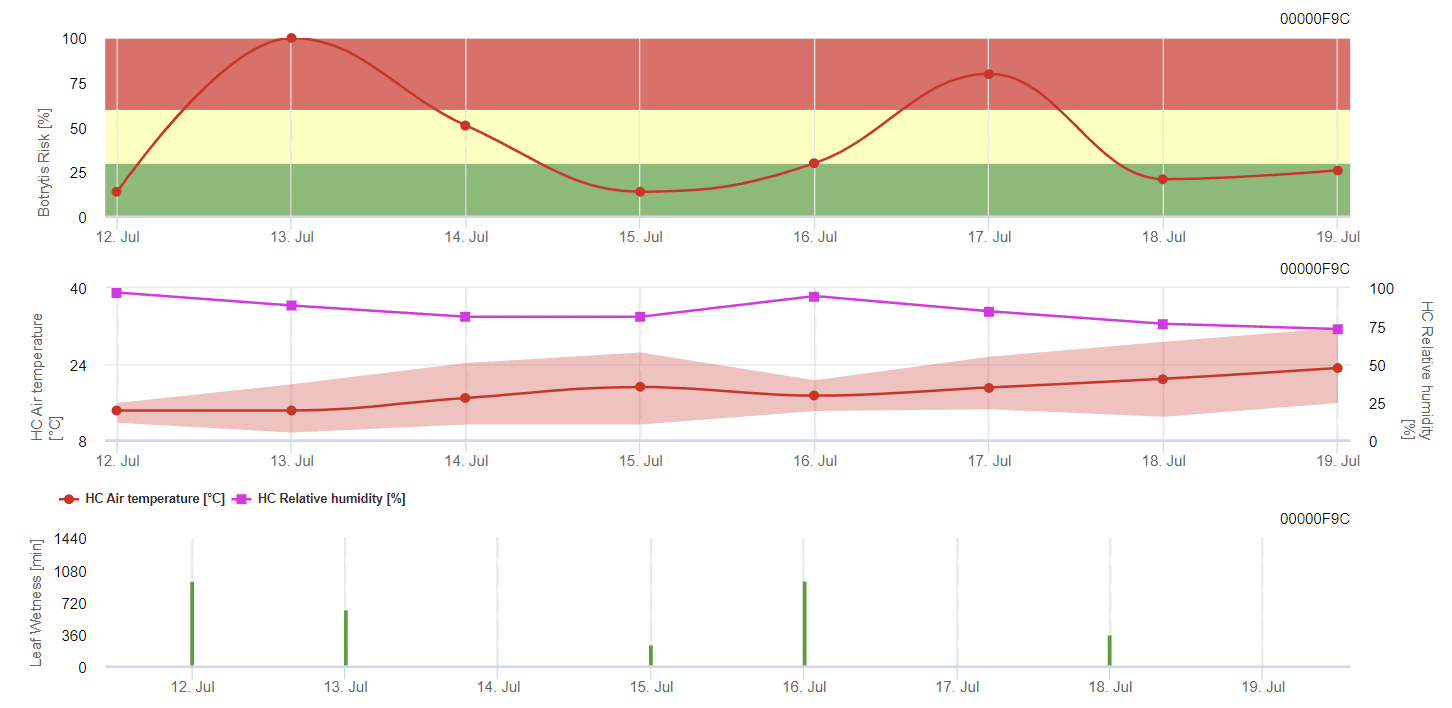
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umiditatea frunzelor
B. cinerea este legată de climatul umed. Ciuperca necesită o umiditate relativă ridicată sau prezența apei libere pentru infectare. Perioadele călduroase cu perioade îndelungate de umezeală a frunzelor vor duce la creșterea riscului de B. cinerea, în timp ce perioadele uscate îl vor reduce. Infecția are loc pe lăstarii tineri în timpul perioadelor umede lungi sau al furtunilor de grindină dăunătoare.
Modelul calculează riscul într-o valoare de la 0 la 100%. Această valoare indică presiunea de B. cinerea la momentul respectiv - dacă avem o valoare de 100%, înseamnă că a existat de mai multe ori o perioadă de umezeală suficient de lungă pentru a infecta țesutul sensibil. Calculăm așa-numitele "puncte umede" între umezeala frunzelor și temperatură cu un maxim inițial de 38400 de puncte (începutul sezonului, care prezintă un risc de 30%). După această perioadă, fiecare perioadă umedă cu aproximativ 4000 de puncte umede crește riscul cu 10% sau, pe de altă parte, fiecare perioadă uscată reduce riscul cu ⅕ din valoarea anterioară.
Literatură
- Broome, J. C., English, J. T., Marois, J. J., Latorre, B. A. și Aviles, J. C. (1995). Elaborarea unui model de infecție pentru putregaiul Botrytis al ciorchinilor de struguri bazat pe durata umezelii și pe temperatură. Phytopathology, 85(1), 97-102.
- Elmer, P. A., & Michailides, T. J. (2007). Epidemiologia Botrytis cinerea în culturile pomicole și viticole. În Botrytis: biologie, patologie și control (pp. 243-272). Dordrecht: Springer Netherlands.
- Williamson, B., Tudzynski, B., Tudzynski, P., & Van Kan, J. A. (2007). Botrytis cinerea: cauza bolii mucegaiului cenușiu. Molecular plant pathology, 8(5), 561-580.
- Ciliberti, N., Fermaud, M., Roudet, J. și Rossi, V. (2015). Condițiile de mediu afectează infecția cu Botrytis cinerea a boabelor mature de struguri mai mult decât tulpina sau genotipul transposonului. Phytopathology, 105(8), 1090-1096.
Agenți patogeni
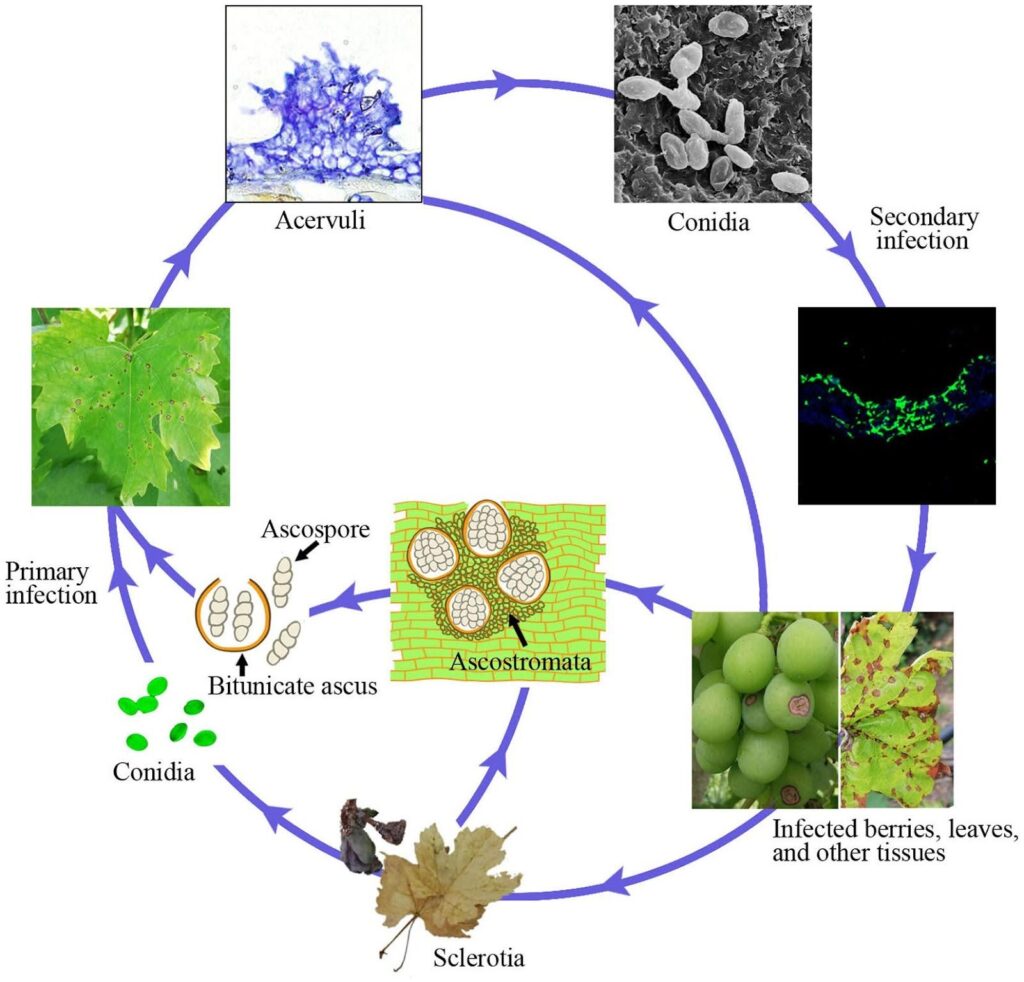
Antracnoza la vița de vie este cauzată de agentul patogen Elsinoe ampelina.
Tulpinile infectate sunt principala sursă a bolii. Scleroții și miceliile care supraviețuiesc în leziuni și boabe în timpul iernii devin activi primăvara și produc ascospori (spori sexuali) și conidii (spori asexuali) în condiții de umiditate (ploaie sau rouă timp de 24 de ore) în intervalul de temperatură 2~40°C.
Acești spori sunt dispersați în noile țesuturi de ploaie, stropi și vânt și, odată ajunși la sol, germinează și formează tuburi germinative și appressoria, inițiind o nouă infecție. Acestea pot infecta frunzele noi, lăstarii, vreascurile și boabele tinere. Vremea caldă reduce durata umedă necesară pentru infecția inițială și perioada de incubare. Germinarea optimă a sporilor are loc la 25~30°C, fiind necesare minimum 3~4 ore de umezeală a frunzelor. La infecții la temperaturi de aproximativ 10°C, perioada de incubare este de aproximativ 14 zile.
Pe măsură ce colonizarea progresează, apar acervuli și se produc noi conidii, care servesc drept inocul de infecție secundară. Acestea sunt responsabile de infectarea rapidă și ulterioară în sezon.
Simptomele
E. ampelina atacă părțile aeriene, suculente ale viței de vie, inclusiv lăstarii, frunzele, pețiolii, vreascurile, ramurile și boabele; leziunile pe lăstari și boabe sunt cele mai frecvente. Strugurii au o rezistență ontogenică la ciupercă, astfel încât țesuturile tinere sunt mai sensibile.
La început apar pete mici maro-roșcate, care se măresc pe vreme umedă, devenind ușor adâncite, cu un centru gri înconjurat de o margine închisă la culoare. Uneori, simptomele pot semăna cu răni provocate de grindină. Centrul necrotic de pe frunze cade de obicei, creând un aspect de "gaură de glonț".
În cazul unei infecții grave, plantele pot prezenta defoliere timpurie, restrângerea creșterii trestiei, ruperea tulpinii, căderea boabelor și întârzierea dezvoltării și coacerii boabelor.
Model FieldClimate
Model de antracnoză în viticultură
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umiditatea frunzelor
- Precipitații
Modelul calculează riscul de antracnoză luând în considerare temperatura aerului, umiditatea relativă, umiditatea frunzelor și precipitațiile.
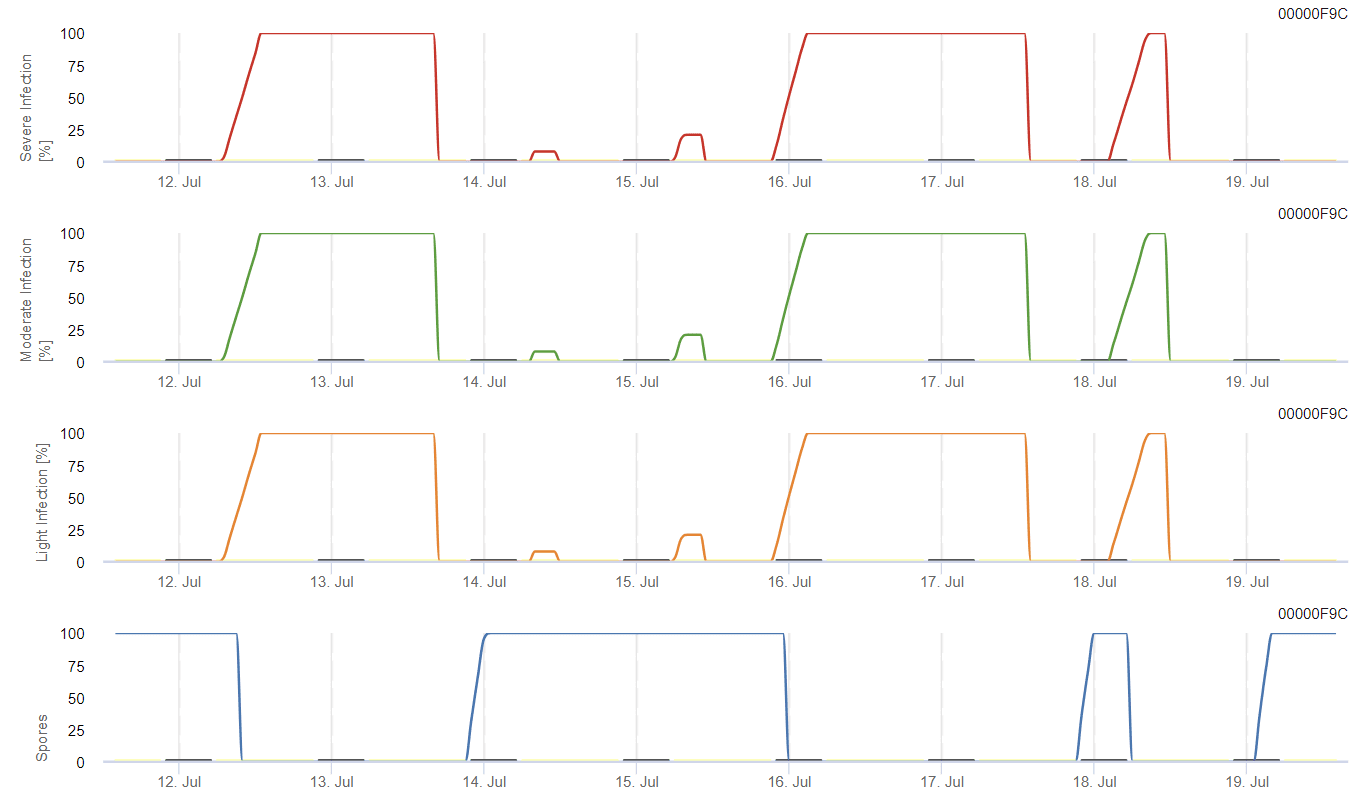
Evoluția posibilă a unei infecții slabe, moderate și severe este prezentată în grafice separate. Se presupune că o infecție este completă atunci când curba crescătoare ajunge la 100%. Ultimul grafic arată sporularea E. ampelina; dacă valoarea ajunge la 100%, se presupune că sporii de E. ampelina sunt prezente.
Pentru ca sporii care iernează să se dezvolte, condițiile trebuie să îndeplinească un interval de temperatură cuprins între 2 și 40°C, o umiditate relativă de peste 90% sau umezeala frunzelor. Odată ce dezvoltarea sporilor atinge 100%, infecția începe să fie calculată. Severitatea infecției depinde de condițiile de umiditate (evenimente de ploaie). Cu toate acestea, dacă umiditatea scade sub 50%, atât dezvoltarea sporilor, cât și infecția încetează să fie calculate.
Literatură
- Li, Z., Dos Santos, R. F., Gao, L., Chang, P. și Wang, X. (2021). Starea actuală și perspectivele viitoare ale antracnozei viței de vie cauzate de Elsinoe ampelina: O boală importantă în regiunile umede de cultivare a strugurilor. Molecular Plant Pathology, 22(8), 899-910
Agenți patogeni
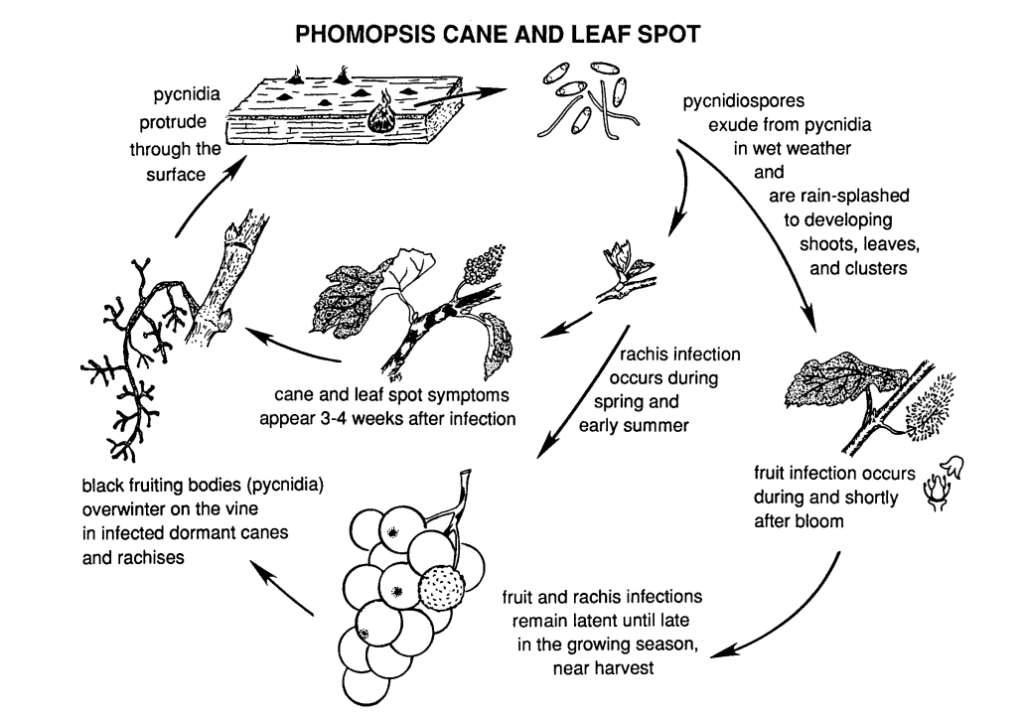
Phomopsis cane and leaf spot in grapevine is caused by Phomopsis viticola.
Ciuperca iernează în părțile lemnoase ale viței de vie și redevine activă în primăvara următoare. Odată ce temperaturile cresc și încep ploile de primăvară, sporii sunt eliberați din structurile de iernare numite picnidii. Pe măsură ce sporii sunt dispersați prin ploaie și vânt pe noi țesuturi, infecția se răspândește.
Perioada de infecție primară coincide de obicei cu ploile de primăvară timpurii, începând la scurt timp după dezmugurire, când lăstarii prezintă un stadiu incipient de creștere. Lăstarii tineri, vârfurile lăstarilor, ciorchinii de fructe și rahisul sunt toate vulnerabile la infecție. Deși vârfurile lăstarilor pot fi infectate pe tot parcursul sezonului de vegetație, infecțiile sunt mai frecvente de la răsărire până la înflorire. În fructe, agentul patogen poate rămâne latent până la coacerea fructelor. Perioada de incubație durează de obicei între 3 și 4 săptămâni.
Simptomele
Pe frunze și pe pețioluri apar mici pete întunecate cu centrul negru înconjurat de margini galbene, care în cele din urmă se contopesc. Frunzele bazale pot deveni deformate și încrețite și pot să nu ajungă la dimensiunea maximă. Atunci când pețiolii sunt foarte infectați, se îngălbenesc și cad, ducând la căderea frunzelor. Frunzele creează un efect de umbrelă care reduce infecțiile ulterioare cauzate de spori.
Pe lăstari apar pete similare, ceea ce duce la crăparea straturilor epidermice la nivelul zonelor infectate. Pe măsură ce petele se unesc, zonele infectate ale lăstarilor capătă o textură scabroasă. În cazurile grave, lăstarii pot să se atrofieze, să se rupă sau chiar să moară.
Leziunile de pe rahis fac ca acesta să devină înfundat și fragil. După o perioadă de latență în timpul verii, ciuperca se reactivează la începutul toamnei, ducând la putrezirea boabelor și a ciorchinilor. Deși infecțiile fructelor nu sunt extinse în general, pe fructe se formează pete, însoțite adesea de pete negre (picnide) pe pielița boabelor. În condiții de umiditate, masele de spori galbeni se pot scurge din pycnidia, ceea ce duce la decolorarea fructelor, la o textură aspră și la mumificare.
Lemnul infectat prezintă un aspect decolorat în sezonul de repaus. Tulpinile și pintenii grav infectați prezintă decolorări întunecate amestecate cu zone decolorate în scoarță. Pycinidia poate, de asemenea, să străpungă suprafața. Temperaturile scăzute, împreună cu afectarea funcțiilor floemului și xilemului, slăbesc și ucid trestiile, pintenii și mugurii.
Model FieldClimate
Model de infecție cu Phomopsis în viticultură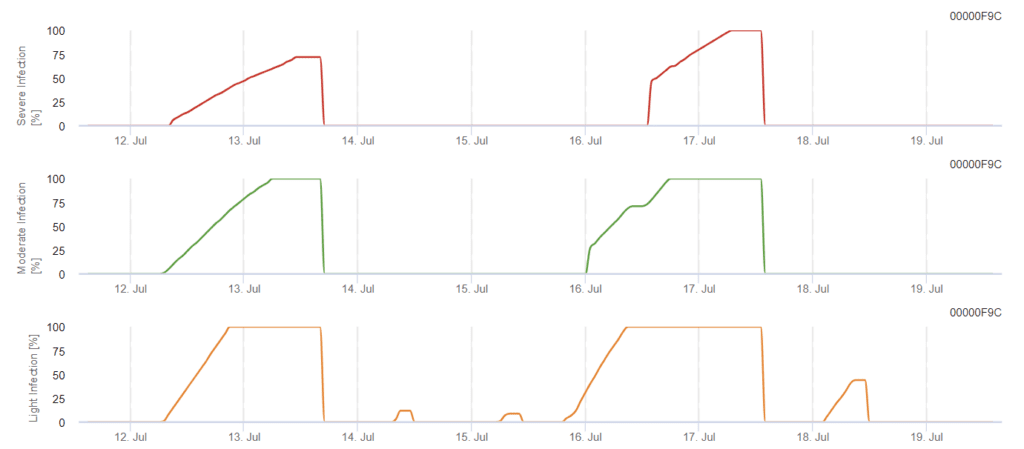
Senzori necesari:
- Temperatura aerului
- Umiditate relativă
- Umiditatea frunzelor
- Precipitații
Modelul determină riscul de infectare cu Phomopsis la temperaturi cuprinse între 5 și 35°C și în condiții de umiditate (frunze umede, umiditate relativă ridicată). Severitatea infecției depinde de cantitatea de ploaie (mai mult de 2 mm), deoarece sporii se distribuie mai rapid către materialul vegetal sănătos. Atunci când apare infecția cu 100%, aceasta indică faptul că pe teren au fost măsurate condițiile optime pentru infectarea țesutului vegetal și, prin urmare, trebuie luate în considerare măsurile de protecție a plantelor.
Literatură
- https://agriculture.vic.gov.au/biosecurity/plant-diseases/grapevine-diseases/phomopsis-cane-and-leaf-spot-of-grapevines
- Bettiga, L. J. (Ed.). (2013). Combaterea dăunătorilor strugurilor (Vol. 3343). Publicații UCANR.
- Pscheidt, J. W., & Pearson, R. C. (1991). Phomopsis cane & leaf spot.