Lucrarea se bazează pe următoarele două articole:
- Boala Black Leaf Streak; Pests and Diseases of American Samoa Number 10, American Samoa Community College Community & Natural Resources Cooperative Research & Extension. 2004
- Sigatoka neagră Un fir în creștere la cultivarea bananelor; Douglas H. Marín, Del Monte Fresh Produce; Ronald A. Romero, Chiquita Brands; Mauricio Guzmán, National Banana Corporation of Costa Rica; Turner B. Sutton, North Carolina State University, Raleigh
Black leaf streak (BLS), sau "Sigatoka neagră", este cea mai dăunătoare boală foliară a bananelor din întreaga lume. Majoritatea zonelor tropicale și subtropicale sunt afectate, cu pierderi raportate de 30-50%. Primul caz de BLS în Pacific a fost semnalat în urmă cu 40 de ani în Valea Sigatoka, Fiji. Majoritatea fungicidelor utilizate în producția de banane sunt destinate combaterii BLS. Banana "Williams" de tip Cavendish, cultivată de majoritatea cultivatorilor comerciali, este foarte sensibilă la această boală fungică. Plantele infectate au mai puține frunze, ceea ce duce la fructe mai puține și mai mici, la o recoltă întârziată, la o coacere prematură și la fructe de calitate inferioară.
Impactul potențial
Sigatoka neagră provoacă distrugerea țesutului frunzelor de bananier, ceea ce afectează capacitățile fotosintetice ale plantei și poate reduce producția cu până la 50%. Sigatoka neagră este unul dintre principalii factori responsabili pentru declinul industriilor de export de banane din țările din Pacificul de Sud. Plantațiile comerciale care produc banane pentru export trebuie să mențină un program costisitor de pulverizare cu fungicide pentru a controla sigatoka neagră și au fost criticate din considerente legate de mediu și de sănătatea umană. Cu toate acestea, dacă nu sunt controlate, fructele produse pe plantele bolnave se pot coace prematur în timpul transportului și pot cauza pierderi suplimentare. Ravagiile bolii au fost controlate prin pulverizări chimice în anii următori (15-17 aplicări de fungicide), dar acest lucru a crescut considerabil costul de producție.
Plante afectate
Simptomele clare ale sigatoka neagră au fost înregistrate doar pe Musa (Banane), Musa paradisiaca (Plantain), Musa acuminata (Banană sălbatică) și Musa acuminata (subsp. bantesii și subsp. zebrina). Soiurile diferă în ceea ce privește reacția lor la agentul patogen. Imunitatea nu este cunoscută și este posibil ca și alte specii și subspecii sălbatice de Musa sunt infectate, dar boala nu se dezvoltă semnificativ. Mycosphaerella fijiensis poate ataca, de asemenea, semințele de banane sălbatice.
Sezonul de apariție
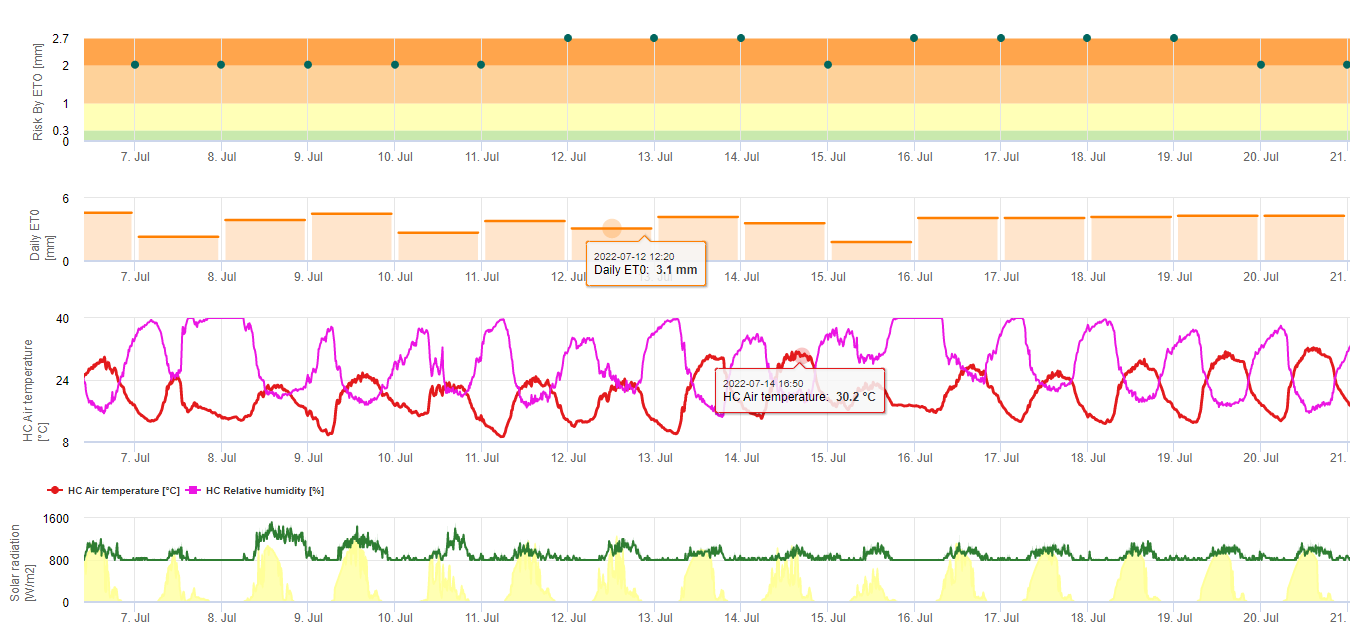
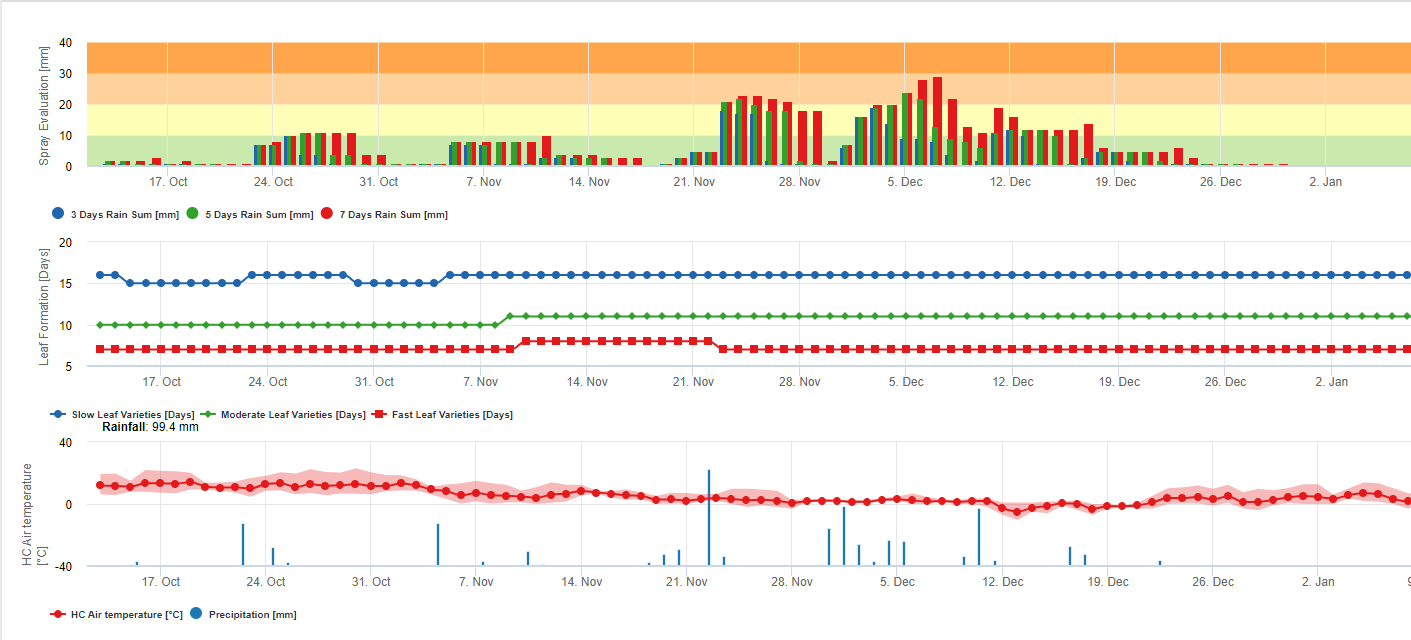
Sporii de sigatoka neagră se formează cu ușurință în timpul verilor tropicale și subtropicale sau în condiții de umiditate ridicată și precipitații, mai ales dacă există o peliculă de apă liberă pe frunze. Principalul mijloc de răspândire este prin spălarea sau împrăștierea sporilor prin ploaie, dar mai târziu, în cursul dezvoltării bolii, sporii sunt, de asemenea, evacuați cu forța în curenții de aer. Infecția se produce pe cele mai tinere frunze ale plantei în timpul și imediat după desfășurare, dar frunzele mai bătrâne sunt mai favorabile dezvoltării bolii. Simptomele vizuale evidente apar la aproximativ 15-20 de zile după infecția inițială.
Simptomele
Primele simptome evidente ale infecției frunzelor sunt linii scurte, de culoare brun-roșiatică, cu o lungime de aproximativ 2 mm. Liniile se transformă în dungi brune de 20-30 mm lungime care se lărgesc în pete eliptice, brune pe fața inferioară a frunzei, negre pe fața superioară. Petele sunt ușor înfundați și sunt înconjurate de un halou gălbui. Apoi, centrul petei mari devine gri, mărginit de o linie neagră și de un halo galben. În cele din urmă, petele se unesc, ucigând frunza.
Epidemiologie
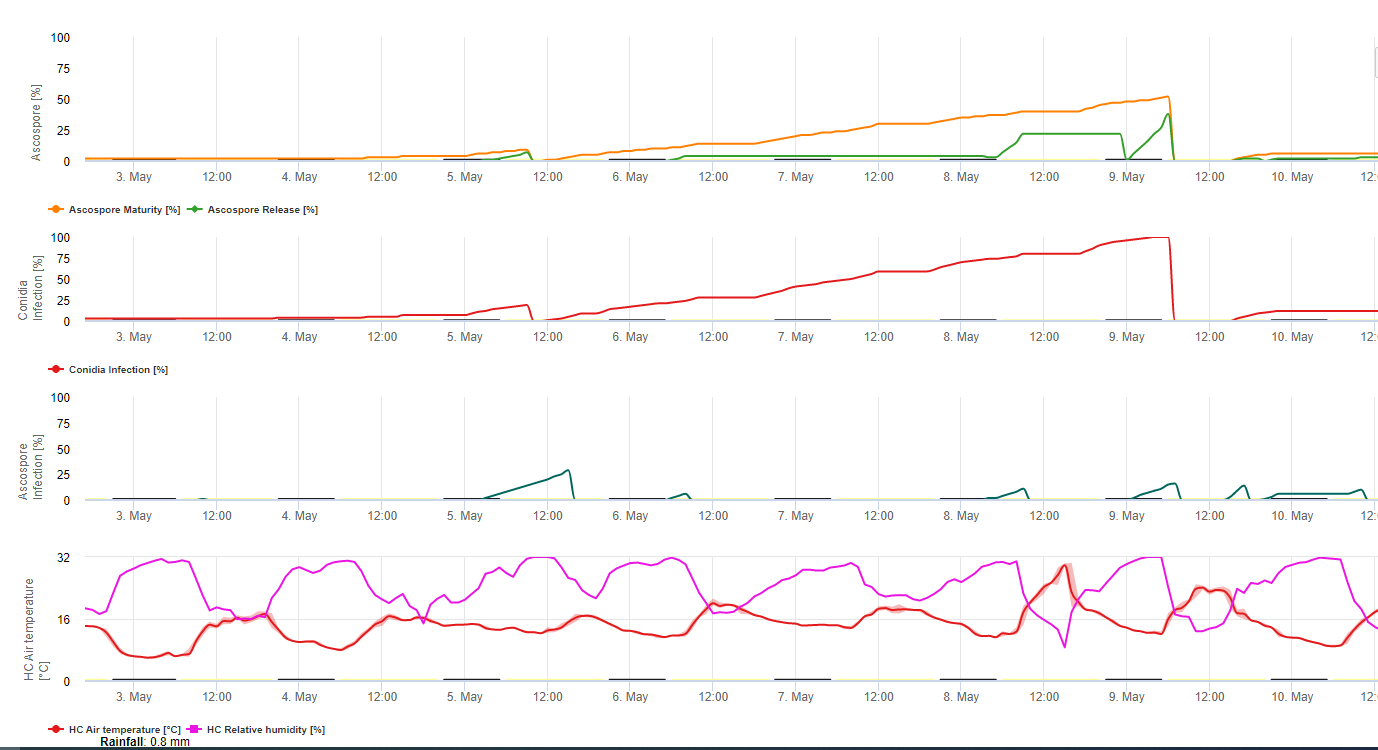
Atât conidiile, cât și ascosporii joacă un rol în răspândirea bolii. Conidiile se formează în condiții de umiditate ridicată, mai ales dacă există o peliculă de apă liberă pe frunze. Acestea se formează în timpul dezvoltării primelor stadii ale bolii (în special în timpul stadiului 2 al liniuței, stadiului 3 al striației și stadiului 4 al petelor). Principalele mijloace de dispersie sunt ploaia și stropirea; conidiile nu sunt detașate de vânt. Conidiile sunt asociate în principal cu răspândirea locală a bolii și sunt importante în perioadele de umiditate ridicată, cu roua abundentă și frecventă și cu ploi intermitente.
Pentru că M. fijiensis produce relativ puține conidii, ascosporii sunt considerați a fi mai importanți în răspândirea Sigatoka neagră (4,20). Cu toate acestea, Jacome et al. (10) au afirmat că conidiile sunt capabile să provoace cantități semnificative de boală, iar simptomele bolii sunt identice cu cele provocate de infecția cu ascospori. El a indicat că conidiile devin mai importante în timpul perioadelor secetoase, când dezvoltarea bolii este întârziată din cauza prezenței unor condiții climatice mai puțin favorabile (10). Ascosporii sunt principalul mijloc de dispersie pe distanțe mai mari în interiorul plantațiilor și în zone noi și reprezintă mijlocul obișnuit de răspândire în timpul perioadelor prelungite de vreme umedă (3,4,6,10, 11,15).
Ascosporii sunt produși în pseudotecii din leziunile mature, care sunt frecvente pe frunzele mai bătrâne ale plantei sau pe frunzele moarte care zac pe sol. Meredith și Lawrence (14) au raportat că pseudotecii sunt produși pe ambele fețe ale suprafeței frunzelor, dar un număr mai mare este prezent pe suprafața adaxială. Dimpotrivă, Gauhl et al. (6) au constatat că pe suprafața abdominală a frunzei se produc mai multe pseudotecii și ascospori. Deși Burt et al. (2) au constatat că aproximativ 4,5 ascospori sunt eliberați pe pseudotecium, acest număr pare foarte mic având în vedere concentrația mare de inocul prezentă în aer în urma ploii (4).
Eliberarea de ascospori
Necesită prezența o peliculă de apă provenită din ploaie sau rouă care îmbibă pseudotecile și are ca rezultat ejectarea forțată a ascosporilor prin stratul limită al frunzei, unde sunt diseminate de curenții de aer (15,20). Maturarea pseudotecii necesită saturație din țesuturile frunzelor moarte timp de aproximativ 48 de ore (3,4,7,10,11,15). În condițiile hawaiiene, concentrațiile de ascospori au crescut în timpul nopții, au fost maxime în jurul orei 0600 și au scăzut semnificativ în timpul zilei. În zilele ploioase, concentrațiile maxime au avut loc la scurt timp după începerea ploii. Creșterile sezoniere ale concentrațiilor medii zilnice de ascospori sunt asociate cu creșterea precipitațiilor și a umidității relative (15). Gauhl (4), lucrând cu banane, a raportat rezultate similare în condițiile din Costa Rica. În timp ce Meredith et al. (15) nu au găsit dovezi că producția sau eliberarea de ascospori a fost afectată de temperaturile minime, Gauhl (4) a determinat că există o reducere a producției de inocul în timpul lunilor mai uscate (sau mai puțin ploioase) ale anului în zona Caraibelor din Costa Rica, care este, de asemenea, sezonul cu cele mai scăzute temperaturi. Ascosporii sunt dispersați de vânt; cu toate acestea, dispersia pe distanțe lungi este limitată la câteva sute de kilometri din cauza sensibilității lor la radiațiile ultraviolete (17). Nu a fost demonstrată o relație consecventă între descărcarea ascosporilor și dezvoltarea bolii, limitând astfel utilitatea captării sporilor pentru prognoza bolilor (5,11). Ascosporii sunt depozitați în principal pe suprafața inferioară a frunzei în timpul desfășurării unei frunze noi, producând un model de infecții în bandă pe partea care este expusă prima dată, ceea ce reflectă depunerea crescută de spori pe frunza cilindrică de candela în timpul desfășurării acesteia, spre deosebire de întreaga lamă deschisă a frunzei. În consecință, cele mai multe infecții apar pe suprafața abdominală a frunzelor (3,7).
Infecție
Tuburile germinative au nevoie de aproximativ 48 până la 72 de ore pentru a pătrunde în stomate (1,20), deși ciuperca poate crește epifită pe suprafața frunzelor timp de până la 6 zile înainte de a pătrunde efectiv în frunze (1). Infecție reușită este promovată de o extindere a perioadele de umiditate ridicată și prezența apei libere pe frunze (3,7,13). Germinarea maximă are loc atunci când există apă liberă. Conidiile germinează într-un interval mai larg de umiditate relativă (92 până la 100%) decât ascosporii (98 până la 100%). Efectul temperatură asupra germinației poate fi caracterizată printr-o funcție de răspuns pătratică, cu un optim estimat de 26,5°C (10). Stover (21) a observat o creștere maximă a tuburilor germinative de ascospori la 26 până la 28°C după 24 de ore de incubație. Jacome și Schuh (8,9) au raportat că frunzele mai bătrâne sunt mai sensibile; cu toate acestea, E. Bureau (comunicare personală) și Romero (18) au observat că frunzele mai tinere sunt mai sensibile (de la prima la a treia) decât cele mai bătrâne.
Perioada de incubație
Timpul dintre infecție și apariția simptomelor variază în funcție de condițiile meteorologice și de sensibilitatea plantelor (68). La bananieri, se estimează că momentul infectării frunzelor coincide cu apariția unei noi frunze din vârful pseudostemului (20). În condiții foarte favorabile în Costa Rica și cu o gazdă susceptibilă, se poate observa că perioada de incubație poate fi de 13-14 zile, în timp ce în perioadele cu vreme nefavorabilă, durata perioadei de incubație se poate prelungi până la 35 de zile. Rapoarte similare există în Nigeria, pe plantain (16). În timpul sezonului ploios, perioada de incubație a fost de 14 zile, dar în sezonul uscat a fost de 24 de zile. Durata perioadei de incubație variază, de asemenea, în funcție de sensibilitatea cultivarului la M. fijiensis. Perioada de incubație a fost de 26,2 și, respectiv, 25,1 zile la soiurile FHIA 1 și FHIA 2, doi hibrizi sintetici tetraploizi dezvoltați de FHIA, în comparație cu 22,4 zile la soiul sensibil Grande Naine (19).
După ce pătrund în frunză, hifele de <M. fijiensis colonizează celulele adiacente timp de aproximativ 7 zile, fără a prezenta semne de perturbare a celulelor. Hifele vegetative pot ieși din stomate și se pot dezvolta pe suprafața frunzei și pot pătrunde în stomatele adiacente sau pot produce conidiofori și conidii. Această creștere epifită permite ciupercii să colonizeze țesutul foliar adiacent, ceea ce duce la dezvoltarea rapidă a simptomelor (4,20). M. fijiensis are o capacitate mai mare de a pătrunde în mai multe stomate decât M. musicola, ceea ce duce la o pătare mai mare decât în cazul primului agent patogen (20).
Perioada latentă
Deși conidiile pot fi produse în leziuni care prezintă simptome timpurii de M. fijiensis și contribuie la epidemie, perioada de latență este definită de momentul în care ciuperca începe să producă leziuni cu pseudotecii și ascospori maturi, care reprezintă principala sursă de inoculum. Ca și perioada de incubație, perioada de latență variază, de asemenea, în funcție de condițiile meteorologice, de sensibilitatea gazdei și de intensitatea infecțiilor. Perioada de latență a variat de la 25 de zile în timpul sezonului ploios (iunie-decembrie) la 70 de zile în timpul sezonului uscat la Guapiles, Costa Rica. Atunci când vremea este foarte favorabilă descărcării ascosporilor și infecției, multe infecții apar pe frunze. Atunci când infecțiile sunt dense, acestea se unesc rapid într-un stadiu foarte timpuriu de dezvoltare, accelerând apariția petelor mature care se caracterizează prin prezența pseudotecilor și a ascosporilor (22). În aceste condiții, frunzele sunt rapid și grav afectate. Perioada de latență variază, de asemenea, în funcție de nivelul de rezistență. De exemplu, timpul de la apariția frunzelor până la apariția primului simptom al petelor mature în aceleași condiții naturale pentru cultivarul Curraré, o banană de gătit aparținând subgrupului plantain, a fost de 44 de zile, comparativ cu 34 de zile pentru cultivarul Valery, o banană aparținând subgrupului Cavendish (4). Termenul "timp de dezvoltare a bolii" este utilizat pe scară largă în literatura de specialitate privind bananele de la tropice pentru a se referi la perioada de latență și este definit ca fiind timpul dintre infecție și formarea petelor mature. Un alt termen frecvent utilizat în literatura de specialitate în domeniul bananierului este "timpul de evoluție a simptomelor" sau "perioada de tranziție", care reprezintă timpul dintre primele simptome și apariția petelor mature (6). Timpul de evoluție a simptomelor oferă o bună indicație a rapidității cu care boala progresează pe frunze.
Literatură:
- Beveraggi, A. 1992. Etude des interactions hôte-parasite chez des bananiers sensibles et resistants inoculés par Cercospora fijiensis responsible de la maladie des raies noires. Teză de al treilea ciclu. Université de Montpellier, Montpellier, Franța.
- Burt, J. P. A., Rosenberg, L. J., Rutter, J., Ramírez, F., și González, O. H. 1999. Prognoza răspândirii pe calea aerului a Mycosphaerella fijiensis, cauza bolii Sigatoka neagră la bananieri: Estimări ale numărului de peritecii și ascospori. Ann. Appl. biol. 135:369-377.
- Fullerton, R. A. 1994. Sigatoka Leaf Diseases.Pages 12-14 in: Compendiu de boli ale fructelor tropicale. R. C. Ploetz, G. A. Zentmyer, W. T. Nishijinia, K. G. Rohrbach și H. D. Ohr, eds. American Phytopathological Society, St. Paul, MN.
- Gauhl, F. 1994. Epidemiologia și ecologia Sigatoka neagră (Mycosphaerella fijiensis Morelet) pe plantain și bananier (Musa spp.) în Costa Rica, America Centrală. INIBAP, Montpellier, Franța.
- Gauhl, F. 2002. Sunt capcanele de spori un instrument util pentru a prezice dezvoltarea bolii Sigatoka în plantațiile comerciale de banane. Pagina 22 în: Int. Workshop Mycosphaerella Leaf Spot Dis. Bananas, 2nd - Programme and Abstracts. (Abstr.)
- Gauhl, F., Pasberg-Gauhl, C., și Jones, D. R. 2000. Ciclul și epidemiologia bolii. Paginile 56-62 în: Boli ale bananului, abacá și enset. D. R. Jones, ed. CAB International, Wallingford, Marea Britanie.
- González, M. 1987. Enfermedades del cultivo del banano. Oficina de Publicaciones de la Universidad de Costa Rica, San José.
- Jacome, L. H., și Schuh, W. 1992. Efectele duratei de umezeală a frunzelor și ale temperaturii asupra dezvoltării bolii Sigatoka neagră la bananierul infectat cu Mycosphaerella fijiensis var. difformis. Phytopathology 82:515-520.
- Jacome, L. H., și Schuh, W. 1993. Efectul temperaturii asupra creșterii și producției de conidiale in vitro și compararea infecției și agresivității in vivo între izolatele de Mycosphaerella fijiensis var. difformis. Trop. Agric. 70:51-59.
- Jacome, L. H., Schuh, W., și Stevenson, R. E. 1991. Efectul temperaturii și al umidității relative asupra germinației și dezvoltării tubului germinativ al Mycosphaerella fijiensis var. difformis. Phytopathology 81:1480-1485.
- Jeger, M. J., Eden-Green, S., Thresh, J. M., Johanson, J. M., Waller, J. M., și Brown, A. E. 1995. Boli ale bananelor. Pagini 317-381 în: Banane și banane verzi. S. Gowen, ed. Chapman & Hall, Londra.
- Marín, D. H., și Romero, R. A. 1992. El combate de la Sigatoka negra. Boletín No. 4, Departamento de Investigaciones, Corporación Bananera Nacional, Costa Rica.
- Mayorga, M. H. 1990. La Raya negra (Mycosphaerella fijiensis Morelet) del plátano y del banano. I. Ciclo de vida del patógeno bajo las conditions del Urabá. Revista ICA 25:69-77.
- Meredith, D. S., și Lawrence, J. S. 1969. Boala neagră a frunzelor de banane (Mycosphaerella fijiensis): Simptomele bolii în Hawaii și note privind starea conidială a ciupercii cauzale. Trans. Br. Mycol. Soc. 52:459-476.
- Meredith, D. S., Lawrence, J. S., și Firman,I. D. 1973. Eliberarea și dispersia ascosporilor la boala neagră a frunzelor de bananier (Mycosphaerella fijiensis). Trans. Br. Mycol. Soc. 60:547-554.
- Mobambo, K. N., Gauhl, F., Pasberg-Gauhl, C., și Zuofa, K. 1996. Evaluarea efectului sezonului și a vârstei plantei asupra plantainului pentru răspunsul la boala Sigatoka neagră. Crop Prot. 15:609- 614.
- Parnell, M., Burt, P. J. A., și Wilson, K. 1998. Influența expunerii la radiații ultraviolete în lumina solară simulată asupra ascosporilor care provoacă boala Sigatoka neagră a bananului și a bananei de câmpie. Int. J. Biometeorol. 42:22-27.
- Romero, R. A. 1995. Dinamica populațiilor de Mycosphaerella fijiensis rezistente la fungicide și epidemiologia Sigatoka neagră a bananelor. Teză de doctorat. North Carolina State, Raleigh.
- Romero, R. A., și Sutton, T. B. 1997. Reacția a patru genotipuri de Musa la trei temperaturi la izolate de Mycosphaerella fijiensis din diferite regiuni. Plant Dis. 81:1139-1142.
- Stover, R. H. 1980. Sigatoka leaf spot of bananas and plantains. Plant Dis. 64:750-756.
- Stover, R. H. 1983. Efectul temperaturii asupra creșterii tubului germinativ al ascosporilor de Mycosphaerella musicola și Mycosphaerela fijiensis var. difformis. Fructe 38:625-628.