

Patogen
Pyricularia grisea, známý také jako Magnaporthe oryzae, je houbový patogen zodpovědný za blast pšenice, významnou chorobu postihující pšenici. Patogen se rozmnožuje především nepohlavně prostřednictvím konidií (nepohlavních spor), které slouží jako primární inokulum pro iniciaci infekce. Tyto konidie se vytvářejí na konidioforech a šíří se větrem, deštěm a mechanickými prostředky, např. zemědělskou technikou. Po dopadu na citlivého hostitele pšenice konidie klíčí a vytvářejí apresoria - specializované struktury, které usnadňují pronikání do rostlinných pletiv. Jakmile se houba dostane dovnitř, kolonizuje buňky hostitele, což vede k rozvoji choroby. Podmínky prostředí hrají zásadní roli v infekčním procesu houby. P. grisea. Houbě se daří v teplém a vlhkém podnebí s teplotami od 15 °C do 30 °C a delšími obdobími vlhka v listech nebo vysoké relativní vlhkosti. K iniciaci infekce je nutná nepřetržitá vlhkost listů po dobu nejméně 14 hodin, takže nadměrná oblačnost, vysoká vlhkost, nadměrné zavlažování a špatně načasovaná závlaha podporují rozvoj choroby. Kromě toho mohou závažnost choroby zhoršit stresové faktory, jako je sucho, zhutnění půdy, nízká výška sečení a nadměrné hnojení dusíkem.
Příznaky
Počáteční příznaky blastózy pšenice se projevují jako vodou nasáklé kosočtverečné léze na listech, které později s postupující chorobou šednou. Tyto léze se mohou rozšiřovat a spojovat, což vede k rozsáhlému napadení listů a nekróze. Napadené klasy vykazují částečné nebo úplné vybělení, které často začíná od černošedého infekčního bodu na řapíku nebo bázi klasů. Při vysokém tlaku inokula se může na jedné lodyze objevit více infekčních bodů, což vede ke značným ztrátám výnosu. Kromě listových příznaků, P. grisea může infikovat další nadzemní části pšenice, včetně kolének a krčků, což vede k hnilobě krčků nebo k výbuchu kolének. To může způsobit zlomení napadených částí, což vede k poléhání a dalšímu snížení výnosu. Schopnost patogenu infikovat různé části rostliny v průběhu vegetace jej činí obzvláště destruktivním, zejména za příznivých podmínek prostředí.
Model FieldClimate
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
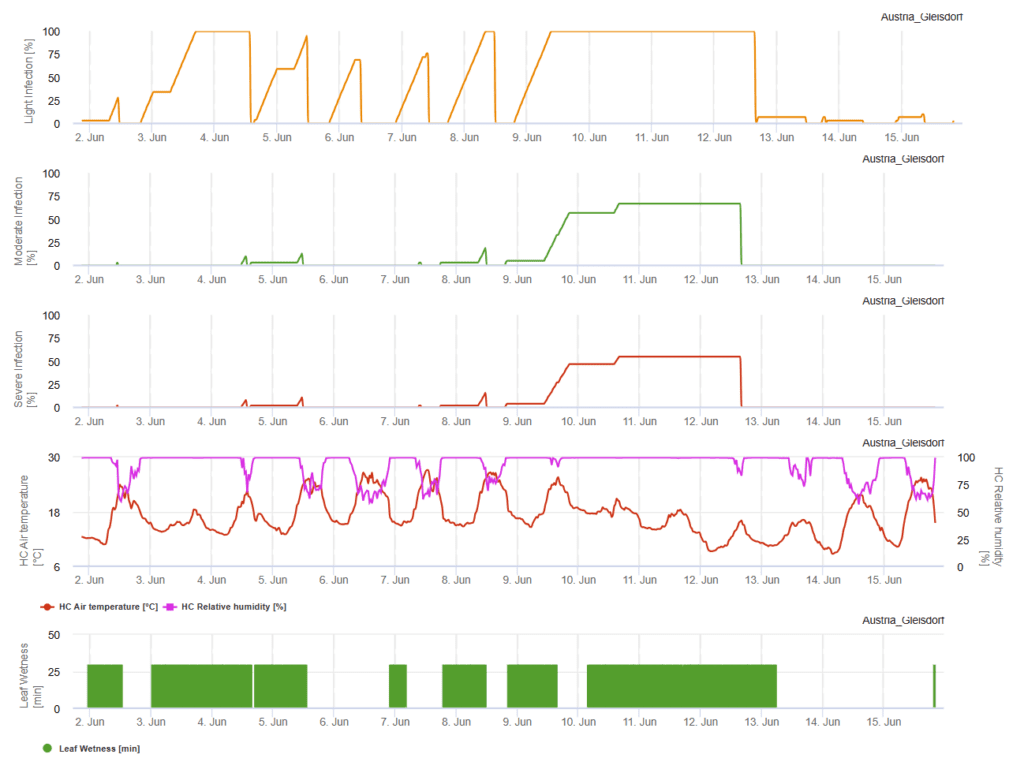
Zatažená obloha, vlhko a mrholení podporují rozvoj a závažnost onemocnění. V FieldClimate počítáme tři třídy závažnosti a při dosažení 100% infekce byly na poli dány optimální podmínky pro houbový patogen. V závislosti na odrůdě, polní historii kurativních opatření na ochranu rostlin je třeba vzít v úvahu, kdy bylo dosaženo 100% lehké, střední nebo silné infekce.
Literatura
- Cruz, C. D., & Valent, B. (2017). Blastická choroba pšenice: nebezpečí v pohybu. Tropická patologie rostlin, 42(3), 210-222.
- Islam, M. T., Croll, D., Gladieux, P., Soanes, D. M., Persoons, A., Bhattacharjee, P., ... & Talbot, N. J. (2016). Výskyt blastózy pšenice v Bangladéši byl způsoben jihoamerickou linií Magnaporthe oryzae. BMC Biology, 14(1), 84.
Patogen
Pyrenophora teres (Drechslera teres) existuje ve dvou formách - P. teres f. teres a P. teres f. maculata které vyvolávají síťovou skvrnu a skvrnu v podobě skvrny.
Askospory uvolněné z pseudotecií infikují zbytky a iniciují primární infekci. Jako primární inokulum slouží v některých případech také mycelia a konidie přenášené semeny. Spory klíčí na listech a pronikají vnější buněčnou stěnou epidermis, takže se patogen může vyvíjet ve velkém intracelulárním měchýřku. Následuje asexuální stadium, během něhož se vytvářejí konidie, které vyvolávají sekundární infekci a zvyšují závažnost onemocnění. Na konci vegetačního období patogen produkuje pseudothecia jako zdroj přesazení a zůstávají na infikovaných zbytcích ječmene.
Příznaky
Příznaky se projevují na listech, stoncích a jádrech, přičemž choroba postupuje od spodní části rostlin k jejich vrcholu. Starší rostliny bývají méně poškozené, protože mají silnější kutikulu, která brání pronikání patogenu, a větší schopnost produkovat antimykotické sloučeniny. Patogeny produkují toxiny, které jsou zodpovědné za příznaky, přispívají k nekróze a chloróze nebo narušují vodní rovnováhu na buněčné úrovni.
V případě síťové skvrnitosti se příznaky projevují jako úzké, tmavě hnědé, podélné léze. Na vysoce odolných odrůdách se vytvoří pouze několik malých tečkovitých lézí bez zřetelného síťovitého vzoru. Naopak u většiny náchylných odrůd se mohou objevit chlorotické nebo vodou nasáklé oblasti obklopující nekrotické léze.
U skvrnitosti se příznaky skládají z tmavohnědých, kruhových nebo eliptických lézí, které jsou často obklopeny chlorotickým nebo nekrotickým halo různé šířky. U méně náchylných odrůd jsou léze obvykle menší a mohou postrádat okolní chlorotickou aureolu.
Těžké infekce mohou vést k úplnému odumření listů, které se pak jeví jako suché. Starší listy obvykle usychají jako první, následované mladšími listy.
Model FieldClimate
Drechslera teres Model
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
- Srážky
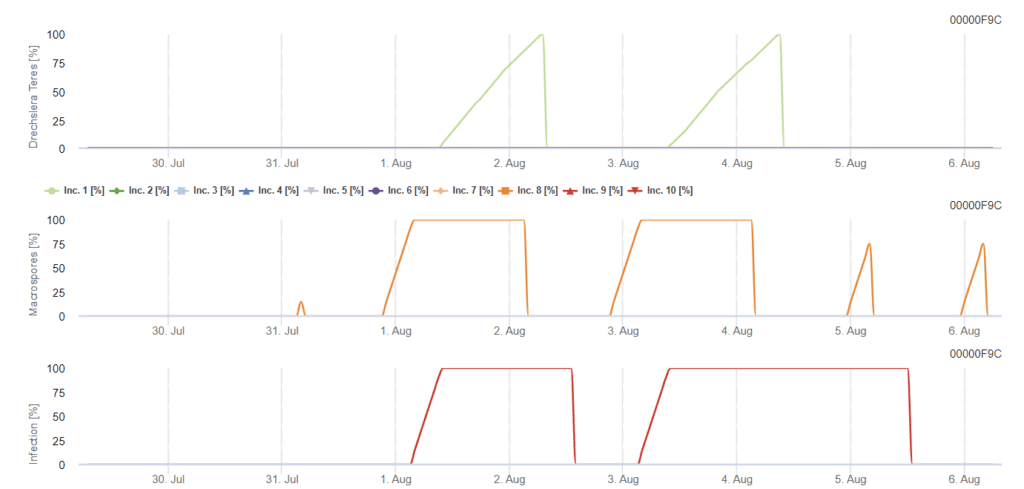
Primární inokulum se vyvíjí při optimálních teplotách mezi 15 °C a 25 °C s optimem 20 °C. Podmínky nad 6 hodin tmy vedou k optimálnímu růstu konidií, jak ukazuje graf sporulace konidií. Pokud sporulační graf dosáhne hodnoty 100%, jsou v terénu nastaveny optimální podmínky.
K dalšímu šíření dochází prostřednictvím větru nebo deště. Infekce rostlinných pletiv vyžaduje vlhké podmínky, jako je vlhkost listů nebo vysoká relativní vlhkost vzduchu po dobu přibližně 10 až 30 hodin při optimálních teplotách 15 °C a 25 °C, jak ukazuje graf infekce. Když dosáhne 100%, je stanoveno, že byly naměřeny optimální podmínky pro infekci na poli. Sluneční světlo může také posílit růst konidioforů, které produkují konidie po změněných podmínkách vlhkých a suchých období s vysokým slunečním zářením a nízkou relativní vlhkostí a vítr podporuje rozvoj choroby.
Literatura
- Backes, A., Guerriero, G., Ait Barka, E., & Jacquard, C. (2021). Pyrenophora teres: taxonomie, morfologie, interakce s ječmenem a způsob kontroly. Frontiers in plant science, 12, 614951.
- Liu, Z., Ellwood, S. R., Oliver, R. P., & Friesen, T. L. (2011). Pyrenophora teres: profil stále škodlivějšího patogenu ječmene. Molekulární patologie rostlin, 12(1), 1-19.
- Obst, A., & Paul, VH (1993). Choroby a škůdci obilovin, Verlag Th. Mann, Gelsenkirchen-Buer.
Patogen
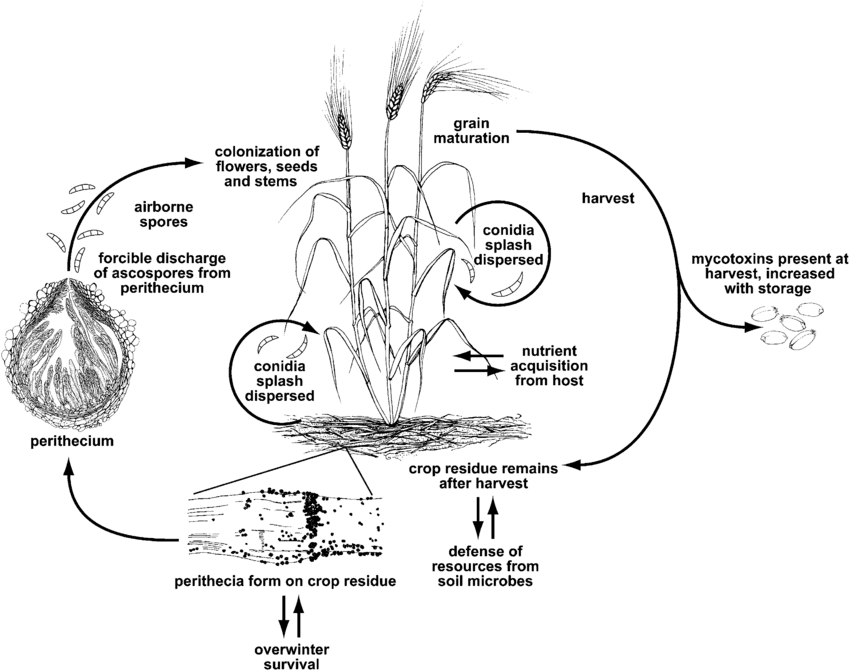
Fusariovou spálu neboli strupovitost způsobuje rod Fusarium. Nejběžnějším druhem je Fusarium graminearum ale nejrozšířenější druhy se mohou lišit v závislosti na regionu.
F. graminearum může infikovat rostliny pšenice v jakékoli růstové fázi. Přežívá zimu na infikovaných rostlinných zbytcích a na jaře dochází k primární infekci, kdy se askospory z perithecií a konidií rozptýlí na pšeničné lusky nebo prašníky. Infikované klásky vytvářejí svraskalá nebo zvadlá zrna, často označovaná jako "náhrobní kameny". I když se zdá, že zrna nejsou zasažena, mohou být kontaminována mykotoxiny. K sekundární infekci dochází prostřednictvím konidií, ale epidemie pšenice závisí především na množství primárního inokula než na sekundárních zdrojích.
Fusariová hniloba vede také k hromadění škodlivých mykotoxinů v plodinách, což představuje potenciální riziko pro zvířata. Tyto mykotoxiny vyřazují obranné mechanismy rostlin nebo chrání houbu před jinými organismy. Hlavním toxinem je deoxynivalenol (DON), který inhibuje syntézu bílkovin a narušuje normální funkci buněk. U lidí, kteří konzumovali pšenici s obsahem DON, se projeví příznaky jako nevolnost, horečka nebo zvracení.
Příznaky

U pšenice a tvrdé pšenice se může objevit jakákoli část nebo celá hlávka vybělená. Částečně bílé a zelené hlávky jsou diagnostickým znakem choroby u pšenice. Houba může napadnout i stonek, což způsobuje hnědé až fialové zbarvení. Za vlhka jsou na napadených kláskách a luscích často vidět růžové až lososově oranžové výtrusy. Mnohá zrna jsou scvrklá a lehká a někdy se jim pro jejich křídový vzhled bez života říká "náhrobní kameny". Za příznivých podmínek se infekce může rozšířit na sousední klásky a infikovat celý klásek včetně řapíku a stopky. Jádra mohou mít matný vzhled a růžové zbarvení v normální velikosti, pokud k infekci došlo v pozdní fázi vývoje.
U ječmene jsou napadené klásky vybělené a mají vzhled nasáklý vodou. Zrna mohou rovněž vykazovat změnu barvy a příležitostně lze za vlhka na kláskách a klasech pozorovat lososově oranžové výtrusy.
Modely FieldClimate
Odhady úrovně FHB na poli jsou založeny na počtech napadených hlávek nebo jader. Barevná vizuální stupnice pro odhad intenzity výskytu FHB v pšenici je k dispozici v NDSU Extension Service (publikace PP-1095): https://library.ndsu.edu/ir/bitstream/handle/10365/9187/PP1095_1998.pdf?sequence=1&isAllowed=y.
Model infekce fuzariózami (riziko fuzarióz)
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
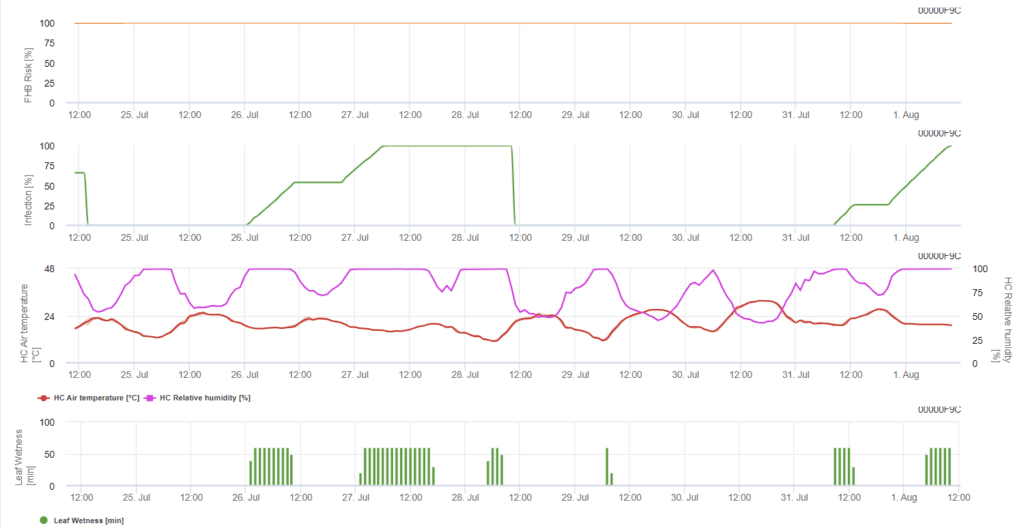
Model určuje riziková období pro infekci. Kdykoli je dosaženo infekce 100%, je riziko velmi vysoké a podmínky pro houbu byly pro infekci příznivé. Znalosti zemědělce o vývojové fázi různých odrůd pšenice dávají možnost rozhodnout o aplikaci kurativního postřiku bezprostředně po infekci.
Původcům fuzarióz na pšenici svědčí vysoké teploty od 20 °C do 30 °C a dlouhá vlhká období. Několikadenní období vlhka na listech vede k časným viditelným příznakům. Příznaky se mohou projevit také po dlouhém období latence, pokud po infekci následuje 18hodinové nebo ještě kratší období vlhnutí listů a po umělém inokulaci při teplotách 15 °C.
Shrneme-li všechny různé kombinace teplot a vlhkosti, které jsme našli v literatuře, rozhodli jsme se upozornit na infekci fuzariózní padlím, pokud teplota a období vlhkosti listů nebo období s relativní vlhkostí vyšší než 85% překročí hodnoty znázorněné v následujícím grafu. Výpočet hodnoty průběhu se řídí vztahem mezi délkou trvání vlhkých podmínek a teplotou.
Model výstrahy před fusariovými mykotoxiny (fusariová hniloba)
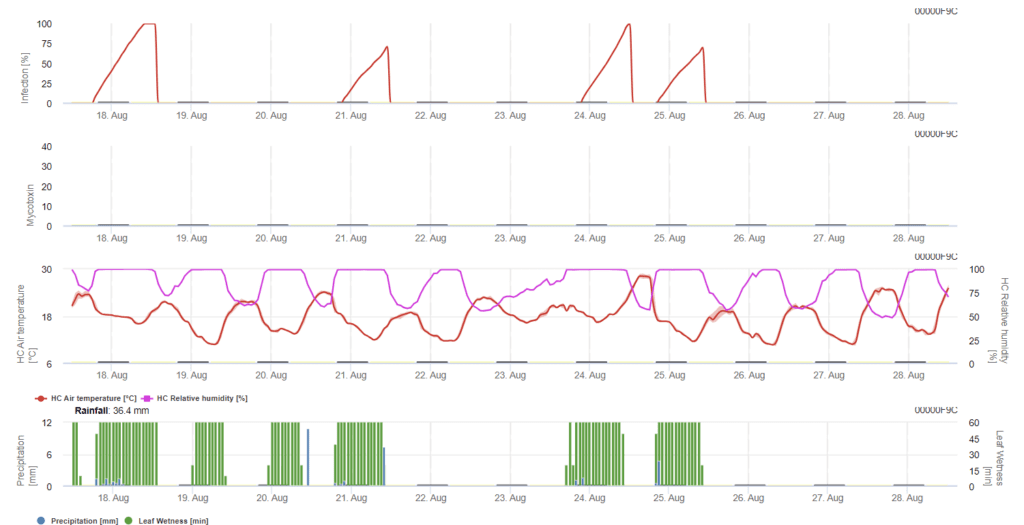
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
- Srážky
v tomto modelu se infekce FHB počítá na základě srážek (potřebné 2 mm), relativní vlhkosti (nad 85%) nebo vlhkosti listů, teploty během procesu. Pokud infekce dosáhne 100%, bylo dosaženo optimálních podmínek pro houbový patogen. Dále model stanoví riziko mykotoxinů FHB.
Pokusy s delšími obdobími vlhnutí listů při infekci fusariovou spále vedly k vysokému obsahu mykotoxinů. Na základě těchto informací se předpokládá, že doba vlhnutí listů 48 hodin nebo více během stadií 61 a 69 představuje vysoké riziko výskytu mykotoxinů.
Zkušenosti získané v průběhu analýzy DON u komerčně pěstované pšenice ukázaly, že dostatečně dlouhé období vlhkých listů pro infekci po počáteční infekci ve fázích 61 až 69 může zvýšit hodnoty DON. V případě delšího období vlhkosti listů mohou mykotoxiny narůstat až do stadia 85.
- BBCH etapa 61: Začátek kvetení; viditelné první prašníky
- Fáze BBCH 69: Konec kvetení; všechny klásky odkvetly, ale některé dehydratované prašníky mohou zůstat.
- BBCH etapa 85: V polovině zrání; měkké těsto s měkkým, ale suchým obsahem zrn, otisk nehtu se neudrží.
Model kumuluje údaj o riziku úměrný průběhu infekce pro každé úspěšné infekční období za zvolené období pro výpočet. Šest dokončených infekcí by vedlo k riziku 100%. Za normálních okolností je období vlhnutí listů vedoucí k fuzariové infekci delší než potřebné minimum. Proto většina fusariových infekcí povede ke zvýšení rizika o více než 17%.
Hodnota rizika mykotoxinů v polní historii. Pšenice pěstovaná po neorané pšenici může nést jen malé riziko, pokud není postřik prováděn v optimální situaci. U nepostřikované pšenice musíme očekávat zvýšené hodnoty DON po 35% rizika. Pšenice po nezpracované pšenici následující po jakékoli jiné plodině může nést vyšší riziko 50%. Pokud máme pšenici po pšenici s orbou, může se riziko zvýšit až na 70%. Pokud riziko dosáhne 100%, měla by být pšenice prvního ročníku testována na DON.
Literatura
- https://www.fao.org/4/y4011e/y4011e0j.htm
- https://scabusa.org/pdfs/NDSU_PP-804_FHB-Small-Grains.pdf
- Lancashire, P. D., Bleiholder, H., Boom, T. V. D., Langelüddeke, P., Stauss, R., Weber, E., & Witzenberger, A. (1991). Jednotný desetinný kód pro růstové fáze plodin a plevelů. Annals of applied Biology, 119(3), 561-601.
- Trail, F. (2009). Pro zkažené vlny obilí: Fusarium graminearum v postgenomické éře. Fyziologie rostlin, 149(1), 103-110.
- Schumann, G. L. (2010). Instruktor zdraví rostlin| Ročník: 10| Rok: 2010| Typ článku: Typ: Plány výuky. Zdraví rostlin, 10.
Patogen
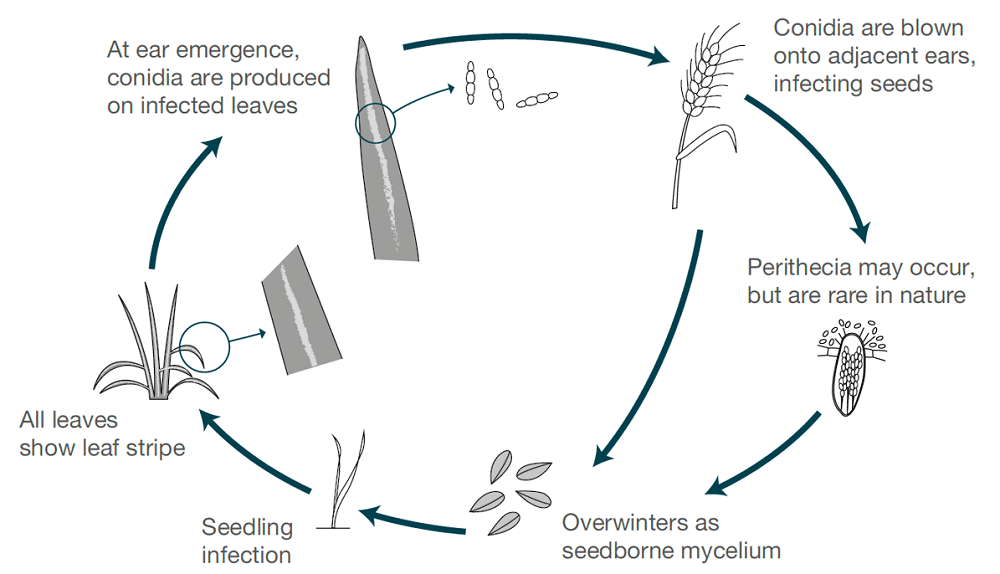
Pruhování ječmene je způsobeno Pyrenophora graminea (Drechslera graminea), patogen přenášený semeny, který přežívá v infikované epidermis semen. Houba infikuje semenáčky v chladných a vlhkých podmínkách. Houba roste v rostlině systémově, infikuje celou rostlinu, produkuje toxiny, ničí buňky a mění barvu listové tkáně mezi žilkami, čímž způsobuje pruhované léze. Pruhování se vyskytuje častěji u ozimého ječmene než u ječmene jarního.
Za vlhkých nebo mokrých podmínek se při rozkvětu zdravých rostlinných výhonků na povrchu listů vytvářejí spory. Spory jsou větrem roznášeny na tyto vyvíjející se klásky, klíčí a způsobují infekce. Jádra jsou nejnáchylnější během raného vývoje. V každé sezóně dochází pouze k jednomu cyklu infekce a produkce spor.
Příznaky

Příznaky jsou nejčastěji patrné ve fázi výskytu v uchu.
V bazální části listu a listové pochvy se zpočátku objevují světle zelené až žluté pruhy. Tyto pruhy postupně hnědnou až tmavě hnědnou, následuje zasychání a rozdělení listové čepele. Rozšiřují se na celou délku listu, nekrotizují a spojují se, což nakonec vede k odumření rostliny.
Rostliny se mohou jevit zakrslé, vytvářet málo klíčků a v závažných případech nevytvářet hlávky ani semena. Délka klasu může být také zkrácena v důsledku špatně vyvinutých hnědých zrn.
Model FieldClimate
Pyrenophora graminea Model
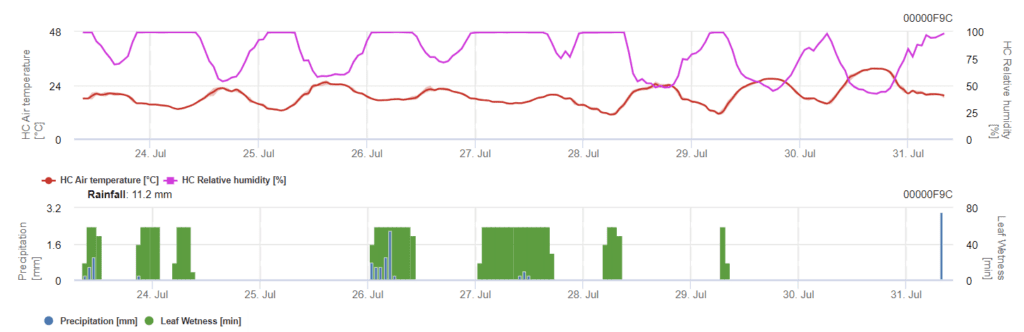
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
- Srážky
Model má dvě fáze. První etapou jsou riziková období v havarijních obdobích elektrárny, rané fáze. Druhá fáze je v době kvetení a vývoje hlávek ječmene. Když infekce dosáhne 100%, byly na poli vytvořeny optimální podmínky pro infekci rostlinných pletiv a systematický růst houby uvnitř.
Literatura
- Richardson, M. J. (1996). Mykologie semen. Mykologický výzkum, 100(4), 385-392. https://ahdb.org.uk/knowledge-library/barley-leaf-stripe-life-cycle-and-disease-symptoms https://plantwiseplusknowledgebank.org/doi/full/10.1079/pwkb.species.46115
Patogen
Existují dvě hlavní septoriové choroby pšenice - septoriová skvrnitost způsobená Septoria tritici. Septoria tritici a skvrny Septoria nodorum způsobené bakterií Septoria nodorum.
Septoria tritici přežívá během léta na infikovaných rostlinných zbytcích a na podzim začíná infikovat rostliny pšenice. Houbě se daří v chladných a vlhkých podmínkách. Má dvě fáze - latentní fázi a nekrotickou fázi. Během latentní fáze houba inokuluje nová rostlinná pletiva a pronikáním přes průduchy následuje kolonizace, při níž se mezibuněčně rozrůstá v rostlinných pletivech. S. tritici nepotřebuje žádné fyzické výživné struktury, ale živí se získáváním živin z hostitelských buněk, které ztratily strukturální integritu. Rostliny mohou vykazovat hypersenzitivní reakci a na napadených pletivech se mohou objevit nekrotické léze jako reakce na infekci.
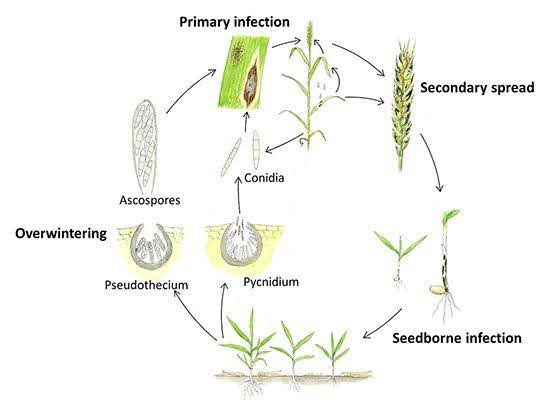
Septoria nodorum přežívá přes léto jako dormantní mycelia, pyknidy a pseudothecia na rostlinných zbytcích. Infekce obvykle začíná na podzim, kdy se pyknidiospory z pyknidií a askospory z pseudotécií rozptýlí větrem a deštěm, což iniciuje primární infekci. Tyto spory pronikají kutikulou listů a po infekci následuje tvorba pyknid. K sekundární infekci dochází při tvorbě pyknidiospor, které se šíří ze spodních listů na horní listy a lodyhy. Patogen rovněž produkuje toxiny, které přispívají k rozvoji choroby.
Příznaky
Skvrnitost Septoria tritici
Na podzim, Septoria tritici příznaky se zpočátku projevují jako malé žluté skvrny na listech, které obsahují drobné černé skvrnky, což jsou plodnice houby. Léze jsou nepravidelného tvaru, od eliptických až po dlouhé a úzké, zvětšují se a v době zralosti hnědnou až červenohnědnou. Choroba obvykle začíná na spodních listech a postupně se přesouvá vzhůru, až nakonec postihne vlajkový list. Za vlhka se houba může rozšířit na hlávky pšenice a způsobit hnědé léze na palicích a oseních, známé jako skvrnitost palic. Septoria tritici blotch může být zaměněna za jiné choroby pšenice. Přítomnost černých houbových tělísek je však klíčovým znakem této choroby. S. tritici. Ačkoli Septoria nodorum může ve vlhkých podmínkách vytvářet také hnědé plodnice, liší se barvou a velikostí, s Septoria tritici plodnice jsou větší.
Skvrna Septoria nodorum
Septoria nodorum příznaky se nejprve objevují hlavně na horních listech. Zpočátku se na listech objevují tmavě hnědé skvrny se žlutou aureolou. Dalším počátečním příznakem je spálení špiček listů. Poškození se rozšiřují a v dospělosti se stávají nekrotickými s tmavě hnědým středem. Uvnitř lézí jsou viditelné drobné tmavě hnědé struktury, které jsou plodnicemi. Konidie uvolněné z plodnic vypadají jako bílé až růžové hmoty. Silně napadené listy mohou být zcela pokryty lézemi, což nakonec vede k odumření listové tkáně. Houba může napadnout také žíně a řapíky: objevují se podobné hnědé léze a choroba postupuje směrem dolů.
Model FieldClimate
Septoriové infekce jsou možné při nízkých teplotách, zatímco teploty pod 7 °C nemusí vést k infekci během 2 dnů. Optimální teploty pro onemocnění je dosaženo v oblasti 16 až 21 °C. Infekce jsou možné v období vysoké relativní vlhkosti vzduchu nebo vlhkosti listů trvajícím 14 hodin nebo déle.
Septoria tritici Model
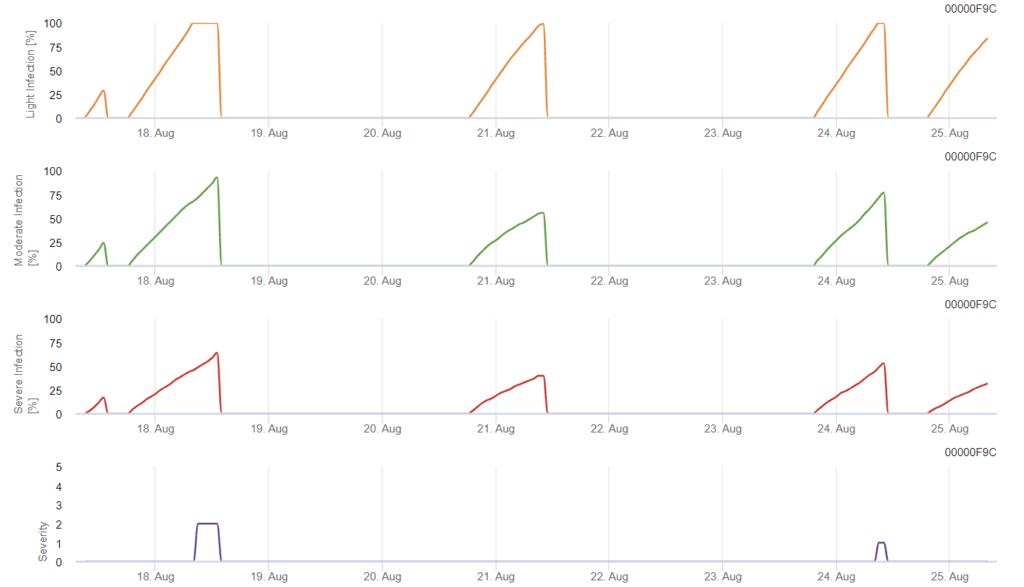
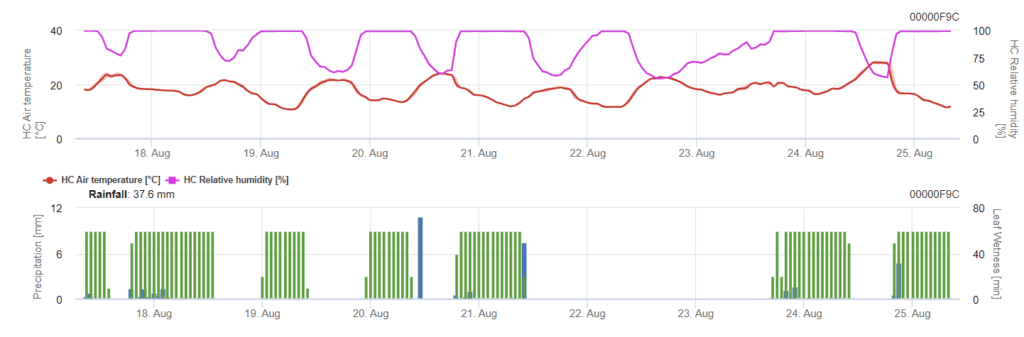
Infekce začíná po dešti o síle 0,5 mm. Optimální teploty pro infekci septoriózou jsou přibližně 16 až 21 °C a vlhkost listů nebo vysoká relativní vlhkost vzduchu po dobu 14 hodin. Septoria tritici se počítá do tří tříd závažnosti. Dále určujeme stupně závažnosti, které podporují hodnocení infekčního tlaku mezi stádii BBCH 10 až 32 pšenice.
- BBCH stadium 10: Vývoj listů; první list přes koleoptile
- Fáze BBCH 32: Prodloužení stonku; uzel 2 nejméně 2 cm nad uzlem 1
- Fáze 51 BBCH: Začátek kvetení s vrcholem květenství, který se vynořil z pochvy a první klásek je právě viditelný.
Rozhodli jsme se nepoužít model pro tvorbu pyknid. Předpokládá se, že podmínkou potřebnou pro tvorbu pyknid je období s relativní vlhkostí vyšší než 85%. Životnost pyknid je 24 hodin. Ve všech klimatických podmínkách, kde má houba šanci se nakazit, najdeme dvě hodiny splňující tuto podmínku téměř každý den kolem východu slunce.
Hodnocení závažnosti infekce
Posoudit Septoria tritici infekčního tlaku mezi stupni 10 a 32 a mezi 32 a 51, musíme posoudit závažnost infekcí na základě klimatických podmínek. Závažnost infekce se hodnotí na stupnici od 1 do 5:
- 1: slabá infekce s deštěm menším než 5 mm
- 2: slabá infekce s deštěm nad 5 mm
- 3: mírná infekce s deštěm menším než 5 mm
- 4: středně těžká infekce s deštěm nad 5 mm/ těžká infekce s deštěm pod 5 mm
- 5: silná infekce s deštěm nad 5 mm
Hodnocení tlaku při onemocnění
Tlak chorob na poli ovlivňují tři faktory - klima, historie pole a náchylnost pěstované odrůdy. Pokud se nám podaří kumulovat hodnoty intenzity choroby od stadia 10 do stadia 32 na hodnotu 4, můžeme očekávat slabý tlak choroby ze strany klimatu. Pokud tato hodnota dosáhne 6, můžeme očekávat mírný tlak choroby, a pokud dosáhne 10, vyšší tlak choroby ze strany klimatu.
Znalost náchylnosti odrůdy a polní historie nás povede k tomu, abychom v dané situaci při slabém nebo mírném tlaku choroby postřikovali nebo ne. Mít kumulativní hodnotu 10 může vést k postřiku ve fázi 32 tak jako tak.
Rozhodnutí o postřiku v pozdější fázi závisí na jarním klimatu. Pokud se nám podaří kumulovat hodnoty závažnosti od fáze 10 do hodnoty 6, můžeme očekávat slabý tlak choroby. Pokud tato hodnota dosáhne 10, můžeme očekávat mírný tlak choroby a pokud tato hodnota dosáhne 15, můžeme očekávat silný tlak choroby vyplývající z klimatické situace.
V systému FieldClimate Septoria tritici závažnost spolu se třemi různými stupni závažnosti infekce. Vzhledem k dešťovým srážkám a dlouhým obdobím vlhka listů byly splněny podmínky pro silnou infekci S. tritici. Stupně závažnosti dosahují nejvyšší hodnoty 5, což znamená, že existuje vysoké riziko infekce.
Septoria nodorum Model
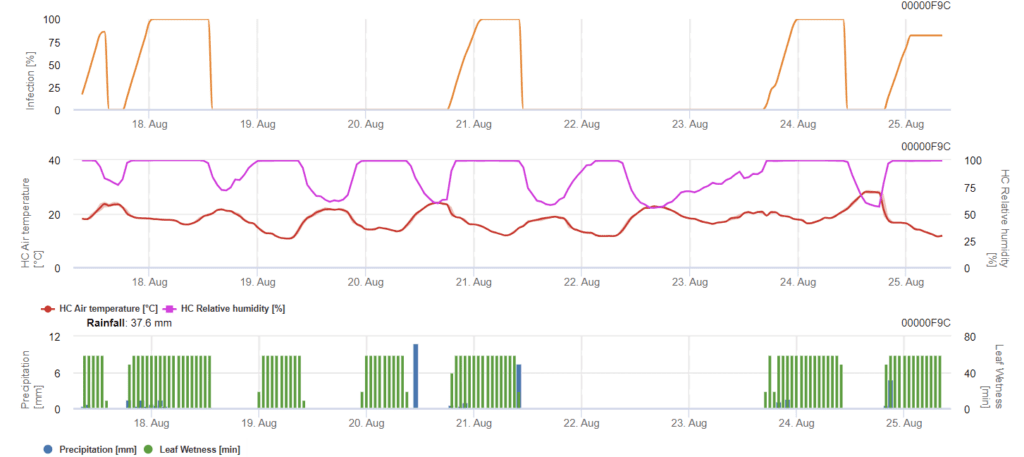
Septoria nodorumse biologie infekce do jisté míry liší od biologie Septoria tritici ale rozdíl není dostatečně velký na to, aby byl vytvořen samostatný model. Proto navrhujeme použít model tritici pro celý komplex onemocnění Septoria. V oblastech s vysokým tlakem S. nodorum, infekce klasifikované jako slabé s hodnotou závažnosti 2 by měly být léčeny vážněji než v jiných oblastech.
Na stránkách Septoria nodorum model vypočítá riziko onemocnění. Pokud riziko dosáhne hodnoty 80%, měla by být zvážena opatření na ochranu rostlin. Pokud riziko dosáhne 100% a infekce je již zjištěna, měla by být provedena systémová opatření na ochranu rostlin (kurativní aplikace).
Literatura
- Brennan, C. J., Benbow, H. R., Mullins, E., & Doohan, F. M. (2019). Přehled známých neznámých v raných stádiích septoriové skvrnitosti pšenice. Patologie rostlin, 68(8), 1427-1438.
- De Wolf, E. D. (2008). Septoria tritici blotch. Mehra, L. K., Adhikari, U., Ojiambo, P. S., & Cowger, C. (2019). Septoria nodorum skvrnitost pšenice. Instruktor zdraví rostlin.
- Solomon, P. S., Lowe, R. G., TAN, K. C., Waters, O. D., & Oliver, R. P. (2006). Stagonospora nodorum: příčina stagonosporové skvrnitosti pšenice. Molekulární patologie rostlin, 7(3), 147-156.
Patogen
Skvrnitost listů způsobuje Rynchosporium secalis.
K primární infekci dochází prostřednictvím askospor nebo konidií na zbytcích plodin. Tyto spory jsou roznášeny větrem a deštěm, klíčí, a když po infekci dojde k rozpadu epidermálních a mezofylových buněk, projeví se příznaky. Sekundární infekce je vyvolána konidiemi z infikovaných listů. Za vlhkých podmínek konidie klíčí na povrchu listů a vytvářejí hyfy, které pronikají kutikulou přímo nad epidermálními buňkami. Následný růst houby je omezen na subkutikulární oblast epidermis.
Příznaky
R. secalis může infikovat kteroukoli část listů. Klíčovým znakem jsou skvrny nepravidelného tvaru a infekce začíná od spodních až po horní listy. Skvrny se zpočátku objevují jako plochy nasáklé vodou. S postupující chorobou jsou uprostřed šedavé v důsledku tvorby spor s hnědým okrajem. Následovat může nekróza a chloróza. Skvrny se spojují a ničí celé listy.
Model FieldClimate
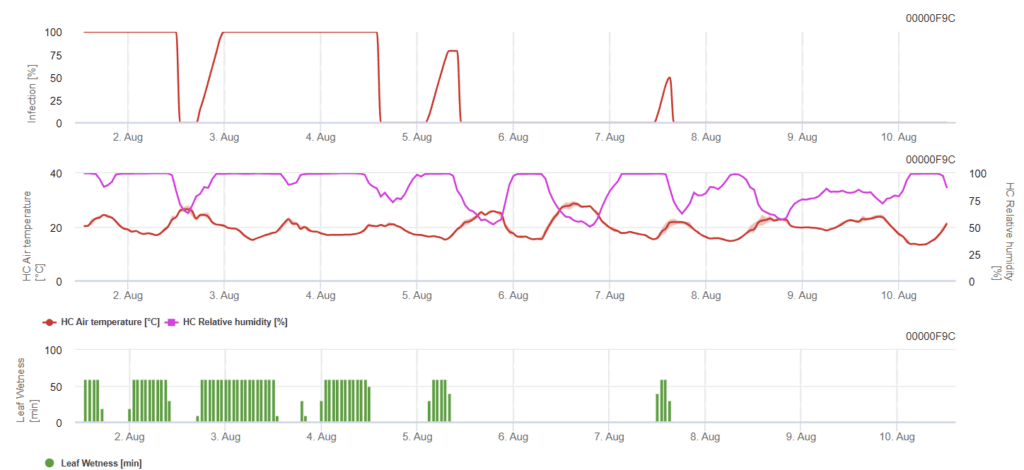
Model listové skvrnitosti Rynchosporium
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
K tomu, aby došlo k infekci, je zapotřebí minimálně 7 až 15 hodin vlhkého olova v závislosti na teplotě (optimální je 20'C). Když graf dosáhne hodnoty 100%, znamená to, že v terénu byly zjištěny optimální podmínky.
Literatura
- Brooks, F. T. (1928). Observations on Rhynchosporium secalis (Oud.) Davis, leaf blotch of barley and rye (Pozorování Rhynchosporium secalis (Oud.) Davis, listové skvrnitosti ječmene a žita). New Phytologist, 27(4), 215-219.
- Fowler, A. M., & Owen, H. (1971). Studies on leaf blotch of barley (Rhynchosporium secalis) [Studie listové skvrnitosti ječmene (Rhynchosporium secalis)]. Transactions of the British Mycological Society, 56(1), 137-152.
- Zhan, J., Fitt, B. D., Pinnschmidt, H. O., Oxley, S. J. P., & Newton, A. C. (2008). Rezistence, epidemiologie a udržitelný management populací Rhynchosporium secalis na ječmeni. Patologie rostlin, 57(1), 1-14.
Patogen
Ramularia collo-cygni, původce listových skvrnitostí rodu Ramularia na ječmeni, se šíří především nepohlavním rozmnožováním prostřednictvím konidií, které pro klíčení a raný vývoj vyžadují vlhkost. Po vyklíčení na povrchu listu houba proniká do listu přes průduchy a mezibuněčně kolonizuje tkáň. Infikované rostliny ječmene nemusí zpočátku vykazovat žádné příznaky, ale poškození se obvykle objeví až po odkvětu.
Životní cyklus R. collo-cygni není zcela objasněna, ale zahrnuje vertikální přenos prostřednictvím infikovaných semen, což umožňuje patogenu přežívat mezi vegetačními obdobími. K sekundárnímu šíření přispívají také konidie přenášené vzduchem, které za příznivých podmínek iniciují nové infekce. Zatímco semena jsou primárním mechanismem přezimování, roli hrají náhradní hostitelé a sekundární houbová struktura, tzv. Asteromella zůstává nejistý a stále se zkoumá.
Příznaky

Příznaky se objevují na obou stranách listů po odkvětu, ale častěji na horních listech. Počáteční příznaky se projevují jako žluté až hnědé skvrny u listových žilek obklopené chlorotickou aureolou. Jakmile se na listech objeví skvrny, listy se stávají chlorotickými a nekrotickými, obvykle začínají od špičky a okrajů listů. Sousední skvrny se mohou spojovat a vytvářet větší tmavé plochy. Lze pozorovat i malé bodové skvrny. Ačkoli příznaky ramuliové skvrnitosti listů mohou být zaměněny s jinými chorobami, klíčovým znakem je, že skvrny na listech jsou ohraničeny na listové žilky.
Pro rozlišení skvrnitosti listů rodu Ramularia existuje tzv. 5R:
- kroužkovaný se žlutým okrajem chlorózy
- Obdélníkový tvar
- Omezeno žilkami listů
- Červenohnědé zbarvení
- Přímo přes list
FieldClimate Model
Ramularia Model
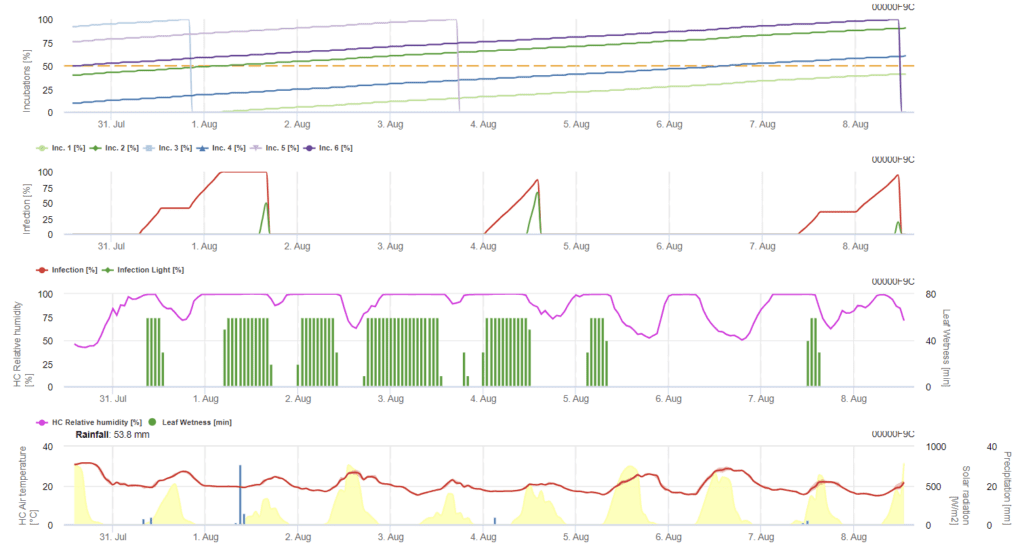
K infekci ječmene ramuliáriemi přispívají především vlhké podmínky. Výpočet tedy začínáme dešťovou událostí, po které následuje vlhkost listů nebo relativní vlhkost vyšší než 85%.
Jsou popsány dva způsoby infekce:
- Infekce: jednou, když vlhké podmínky pouze podporují vznik infekce.
- Infekce: jednou, když je k ukončení infekce potřeba globální záření.
Dále, kdykoli je dosaženo infekce 100%, začne model určovat inkubační dobu. Když inkubace dosáhne 50%, doporučuje se kontrola pole pro nové inokulum, a když dosáhne 100%, předpovídají se nekrotické léze na listech.
Literatura
- Havis, N. D., Brown, J. K., Clemente, G., Frei, P., Jedryczka, M., Kaczmarek, J., ... & Hess, M. (2015). Ramularia collo-cygni-an emerging pathogen of barley crops (Ramularie kolopásná - nově se objevující patogen ječmene). Fytopatologie, 105(7), 895-904.
- Huss, H. (2002, duben). Biologie Ramularia collo-cygni. In Proc. Second Int. Workshop Barley Leaf Blights. Aleppo, Sýrie (s. 321-328).
- Walters, D. R., Havis, N. D., & Oxley, S. J. (2008). Ramularia collo-cygni: biologie nového patogenu ječmene. FEMS Microbiology Letters, 279(1), 1-7. https://grdc.com.au/__data/assets/pdf_file/0025/443509/GRDC_FS2103_Ramularia_03.pdf
Patogen
Rhizoctonia kořenová hniloba pšenice je způsobena Rhizoctonia solani. Patogen může v půdě přežívat mnoho let ve formě sklerocia. V některých případech přežívá také jako mycelia na rostlinných zbytcích. Po zvýšení teploty se sklerocia stávají aktivními a vytvářejí masu hyf, což umožňuje infekci přichycením na kořenech. Mycelium vytváří takzvané "infekční polštáře" a houba napadá a kolonizuje kořenové pletivo. Infekce kořenů oslabuje sazenice, což vede ke špatnému růstu a snižuje schopnost transportu vody a živin do nadzemních rostlinných pletiv, což nakonec vede k úhynu rostlin.
Příznaky
Brzy se objevují holé skvrny o velikosti od několika centimetrů až po několik metrů v průměru. Tyto holé skvrny jsou často spojeny se zakrnělými rostlinami, které mohou vykazovat žloutnutí, vadnutí nebo dokonce fialové zbarvení. Napadené rostliny mohou být silně zakrslé a vykazovat známky stresu ze sucha nebo nedostatku živin, což může v některých případech vést k předčasnému úhynu.
Na stoncích a kořenech pod hranicí půdy vznikají červenohnědé léze, které se s postupující chorobou prohlubují. Kořenová hniloba může omezit nodulaci a způsobit, že se kořenová kůra snadno láme, což vede ke vzniku hnědých špiček kopí.
Model FieldClimate
Model rizika Rhizoctonia solani
Potřebné senzory:
- Teplota vzduchu
- Sluneční záření
- Vlhkost listů
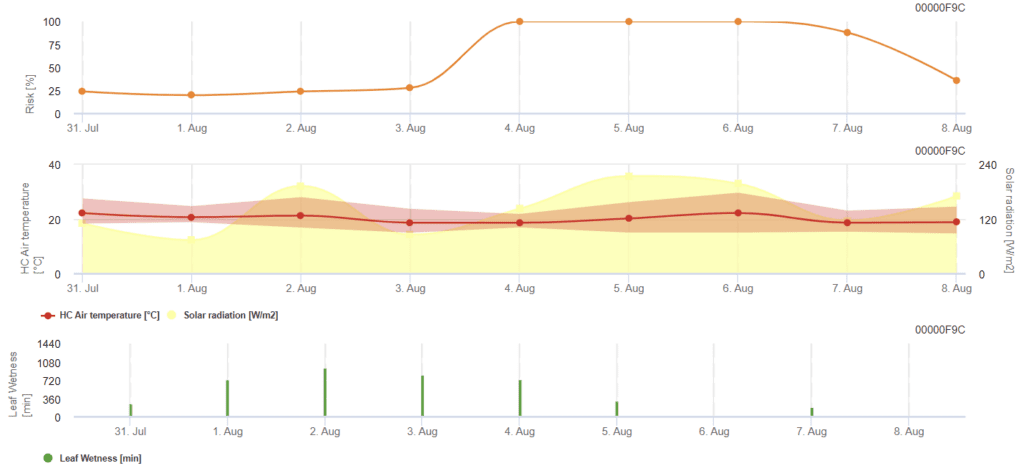
Model vypočítává riziková období Rhizoctonia solani kontrolou podmínek za posledních 120 hodin. Výsledkem je hodnota rizika od 0 do 100, což znamená příznivé podmínky pro onemocnění.
Při nízkém riziku není třeba používat žádné postřiky. V obdobích se středním rizikem může být interval postřiku prodloužen a v obdobích s vysokým rizikem může být nutné jej zkrátit nebo doporučit účinnější použití směsi.
V případě postupného zvlhčení listů kumuluje každou minutu hodnoty závislé na teplotě:
- 12 °C až 15 °C: kumulace 1 za minutu
- 16 °C až 17 °C: kumulace 2 za minutu
- 18 °C a více: kumulace 4 za minutu
Na konci období vlhkosti listů vyhodnotí kumulované hodnoty:
- Hodnoty > 4096: Riziko se zvýší o 64 bodů a od hodnoty se odečte 4096.
- Hodnoty reziduí > 2048: Riziko se zvýší o 16 bodů a od hodnoty se odečte 2048.
- Hodnoty reziduí >1024: Riziko se zvýší o 4 a od hodnoty se odečte 1024.
Pokud je globální záření postupně vyšší než 800 W/m², kumuluje čas v minutách a vyhodnocuje hodnoty, kdy se záření sníží:
- Hodnota > 512: Riziko - 32 bodů, hodnota - 512
- Hodnota > 256: Hodnota - 256, riziko - 8 bodů
- Hodnota > 128: Riziko - 2 body, hodnota - 128
Literatura
- https://cropprotectionnetwork.org/encyclopedia/rhizoctonia-root-rot-of-wheat#:~:text=It%20is%20caused%20by%20Rhizoctonia,result%20in%20premature%20plant%20death.
- https://ahdb.org.uk/knowledge-library/rhizoctonia-stunt-symptoms-and-risk-in-cereals
- https://cropwatch.unl.edu/rhizoctonia-root-rot#:~:text=Disease%20Symptoms&text=These%20reddish%20brown%20lesions%20may,uneven%20because%20of%20stunted%20plants.
- https://extensionaus.com.au/FieldCropDiseasesVic/docs/identification-management-of-field-crop-diseases-in-victoria/soil-borne-diseases/rhizoctonia-root-rot/
- https://www.florimond-desprez.com/es/wp-content/uploads/sites/6/2015/11/rhizoctonia_eng.pdf
Patogen
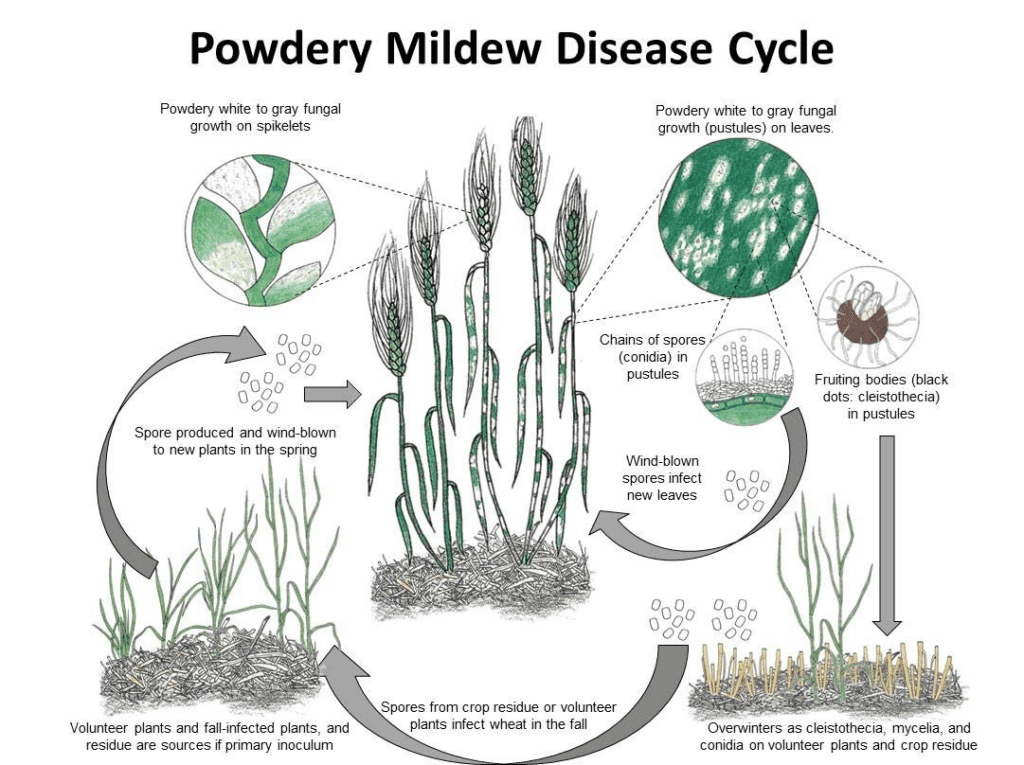
Blumeria graminis je původcem moučnatosti pšenice a ječmene. Může přezimovat ve formě mycelia nebo kleistothecia. Na jaře spory klíčí a infikují hostitelské rostliny za chladných a vlhkých podmínek, aniž by vyžadovaly volnou vodu na povrchu rostlin.
Houba se rozmnožuje nepohlavně i pohlavně.
V asexuálním cyklu prochází patogen rychlými cykly infekce a produkce konidií (asexuálních spor), které jsou klíčové pro šíření nemoci. Konidie se v myceliu za příznivých podmínek tvoří každých 7 až 10 dní. Po rozptýlení větrem tyto konidie dopadnou na nového hostitele, vyklíčí a vytvoří apresoria a haustoria, která z rostliny získávají živiny. Konidie Blumeria graminis mohou klíčit i při nízké vlhkosti a různých teplotách.
V pohlavním cyklu houba vytváří kleistothecia, odolné struktury, které dobře přežívají nepříznivé podmínky. Kleistothecia obsahují askospory (pohlavní spory) a jejich uvolněním dochází k nové infekci.
Příznaky

Prašná plíseň u pšenice a ječmene ovlivňuje výnos zrna tím, že snižuje počet klasů a velikost a hmotnost zrna. Čím dříve k infekci dojde, tím větší jsou škody.
Ječmen vykazuje odolnost vůči padlí v závislosti na věku. Jak ječmen dozrává, mohou se na listech místo typických skvrn od mycelia objevit ostře ohraničené, černohnědé skvrny ("dehtové skvrny").
Pšenice je obzvláště náchylná k moučnivce v období mezi vzcházením a mléčnou zralostí, zejména pokud jsou napadeny praporcovité listy a plevy. Ve srovnání s ječmenem však pšenice snáší ranou infekci lépe.
Na listech, stoncích a hlávkách se objevuje bílošedá moučnatá plíseň. Houba se nejvíce projevuje na horní straně listů, může se však objevit i na spodní straně. Zpočátku se puchýřky jeví jako bílé, ale postupně se mění na šedé až hnědé, jak dozrávají, a mohou se spojovat a vytvářet větší celky často obklopené chlorózou. Na opačné straně infikovaných listů jsou pletiva žlutá, později se zbarvují do hněda nebo hněda. Cleistothecia jsou viditelné i jako malé kulaté černé tečky ve starších šedých koloniích. V závažných případech mohou listy odumírat.
Model FieldClimate
Model prašné plísně pšenice
Potřebné senzory:
- Teplota vzduchu
- Vlhkost listů
- Sluneční záření
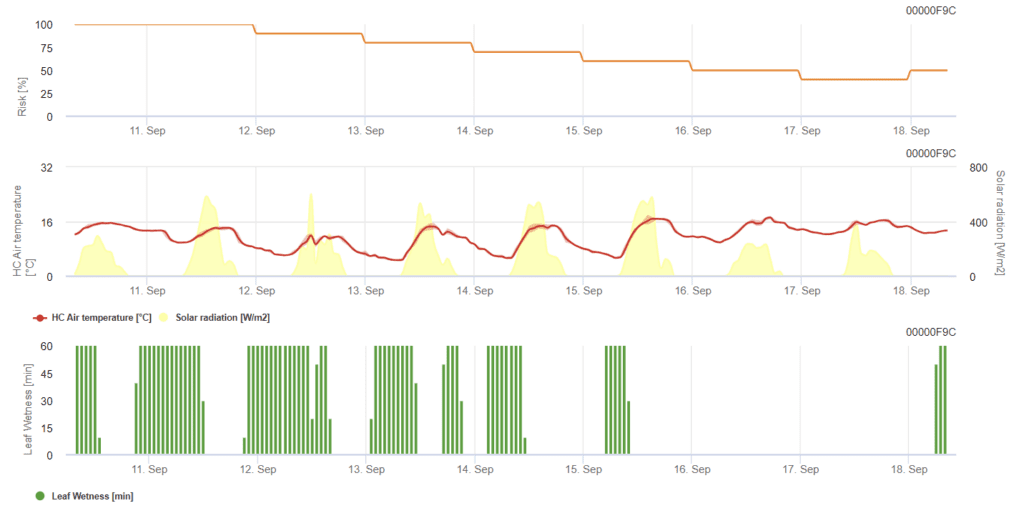
Model určuje riziko výskytu plísně moučnaté na stupnici od 0 (žádná infekce) do 100% (plná infekce). Je relevantní ve fázi BBCH 21 až 39 pro ječmen a 21 až 75 pro pšenici a oves.
Stupeň BBCH je růstová stupnice pro popis vývoje rostlin. Stupnice má dvě číslice a pohybuje se v rozmezí 00 až 99 - 00 se vztahuje k ošetření osiva a 99 znamená ošetření po sklizni. První číslice určuje konkrétní fázi vývoje; 0 je klíčení, 1 je vývoj listů, 2 je obdělávání, 3 je prodlužování stonku, 4 je zakořeňování, 5 je vzcházení květenství a rašení, 6 je kvetení a antecese, 7 je vývoj plodů, 8 je dozrávání a 9 je stárnutí.
- BBCH 21: Kultivace - Začátek kultivace; první kultivace zjistitelná.
- BBCH 39: Prodlužování stonku - stadium praporcového listu; praporcový list je zcela rozvinutý a ligula je právě viditelná.
- BBCH 75: Vývoj plodu - středně mléčný; obsah zrn mléčný a zrna dosahují konečné velikosti, ale jsou stále zelená.
Riziko se zvyšuje při teplotách mezi 12 °C a 21 °C po většinu hodin dne a při nízkém globálním záření. Vlhkost listů, vysoká globální radiace a vysoké teploty nad 32 °C naopak riziko snižují.
Literatura
- Both, M., & Spanu, P. D. (2004). Blumeria graminis f. sp. hordei, obligátní patogen ječmene. Roční přehledy rostlin, 11, 202-218.
- Cunfer, B. M. (2002). Prašná plíseň. Pšenice chlebová: šlechtění a produkce, 30, 317-330.
- Der Gräser, E. M. Krankheiten und Schädlinge des Getreides. Lancashire, P. D., Bleiholder, H., Boom, T. V. D., Langelüddeke, P., Stauss, R., Weber, E., & Witzenberger, A. (1991). Jednotný desetinný kód pro růstové fáze plodin a plevelů. Annals of applied Biology, 119(3), 561-601. https://ohioline.osu.edu/factsheet/plpath-cer-11
Patogen
Existují tři významné rzi a patogeny:
- Rez listová/hnědá: Puccinia triticina
- Stonek/ Černá rez: Puccinia graminis
- Pruh/žlutá rez: Puccinia striiformis
Hnědá rez je nejčastější choroba pšenice. Černá rez je také známá jako rez letní kvůli hojnému výskytu lesklých černých teliospor. Je to nejničivější choroba rzi, která za příznivých podmínek způsobuje 50 % ztrát během jednoho měsíce. Žlutá rez je choroba pšenice v chladnějším podnebí, obecně spojená s vyššími nadmořskými výškami a severními zeměpisnými šířkami. Jejím charakteristickým znakem jsou žlutě zbarvené urediniospory. V závažných případech může způsobit stejně velké škody jako černá rez.
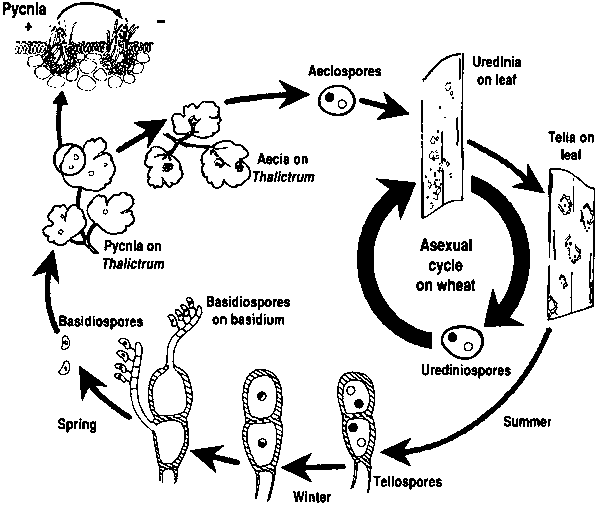
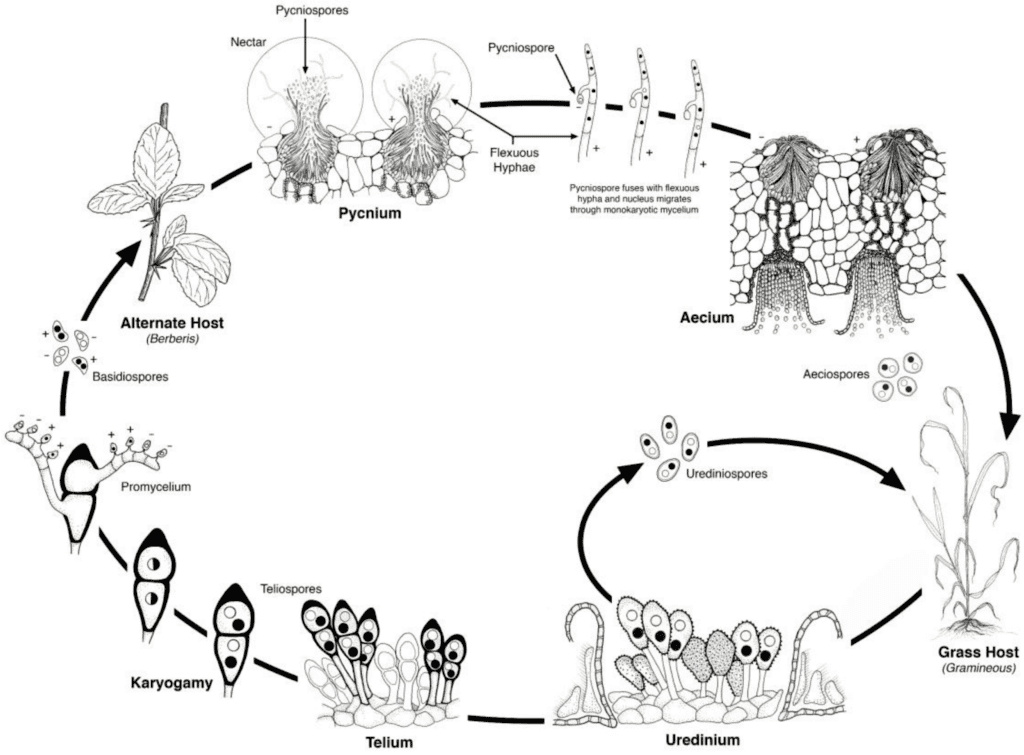
Patogeny rzi pšeničné procházejí asexuálním i sexuálním reprodukčním cyklem a vyžadují dvě hostitelské rostliny - hospodářského hostitele a náhradního hostitele. Hospodářským hostitelem je pšenice a náhradním hostitelem je obvykle plevel nebo původní rostlina, což houbám umožňuje projít pohlavní fází. Puccinia striiformis není známo, že by měl nějakého náhradního hostitele, skládá se z urediniálního a teliálního stadia.
V asexuálním cyklu se urediniospory vytvářejí na hostitelské rostlině a mohou ji opakovaně infikovat, což za příznivých podmínek vede k rychlé a rozsáhlé infekci.
Pohlavní cyklus patogenu začíná, když teliospory, které se vytvoří při urediniální infekci hostitelské rostliny, vyklíčí, projdou meiózou a vytvoří bazidiospory. Tyto bazidiospory se rozptýlí na alternativního hostitele, kde iniciují infekci, což vede k tvorbě pyknií, které obsahují pykniospory a receptivní hyfy. K oplození dochází, když se pykniospory z jednoho typu páření přenesou nektarem na opačný typ. Po oplození se na spodní straně listů alternativního hostitele vytvoří aeciální pohárky, z nichž se uvolňují aeciospory. Tyto aeciospory se rozptýlí a infikují nové hostitele, následuje urediniální infekce, čímž se uzavírá životní cyklus patogena.
Příznaky

A) Puccinia graminis
B) Puccinia striiformis
C) Puccinia triticina
Puccinia triticina
P. triticina napadá především listové čepele, ale příležitostně může napadat i listové pochvy, řapíky a řapíky a vytvářet malé žluté skvrny na horním povrchu listů. Tyto skvrny se s postupující chorobou mění v oranžově zbarvené puchýřky obklopené žlutou aureolou. Z puchýřků se vytvářejí oranžové nebo černé výtrusy viditelné na listech. Na rozdíl od černé rzi vytváří méně teliospor. Na některých místech se mohou objevit přecitlivělé skvrny nebo uredinia obklopená chlorotickými nebo nekrotickými plochami.
Puccinia graminis
P. graminis napadá především stonky, ale může napadat i listy, pochvy, lodyhy, osiny a dokonce i semena. Zpočátku se objevují červenohnědé léze a s postupem choroby se v puchýřcích tvoří černé teliospory. Na stoncích a listových pochvách se nejprve vytvoří masy červeně cihlových urediniospor a objeví se malé chlorotické skvrny, které se vyvinou v lineární nebo kosočtverečné pustuly, které se mohou zvětšovat. V dospělosti se tvorba urediniospór zastaví a vytvoří se vrstva černých teliospor, což způsobí, že stonky koncem sezóny zčernají. V závažných případech infekce oslabuje stonky rostlin a vede k jejich poléhání.
Puccinia striiformis
Charakteristickým rysem P. striiformis je přítomnost rovných žlutých puchýřků. TJey se objevují v úzkých, podlouhlých pruzích různé délky, často doprovázených chlorózou a nekrózou. Jak choroba dozrává, uvnitř puchýřků se vyvíjejí žlutooranžové výtrusy a okolní tkáně hnědnou a vysychají, což vytváří spálený vzhled.
Model FieldClimate
Puccinia Rust Model
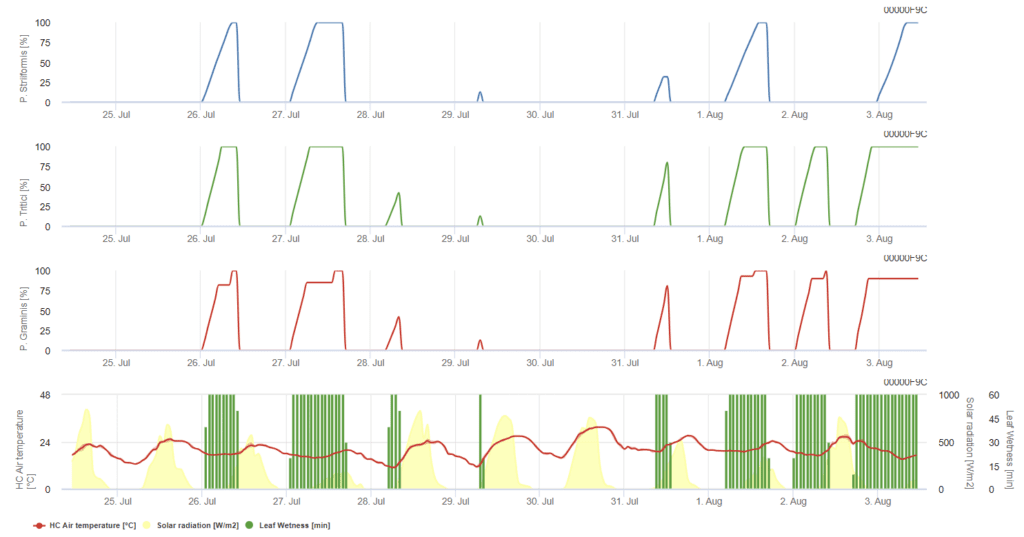
P. triticina, P. graminis, P. striiformis jsou znázorněny ve třech barevných grafech.
Model infekce Puccinia triticna
Model předpokládá, že infekce potřebuje kumulativní hodinovou teplotu 90 °C vlhkosti listů v rozmezí teplot vzduchu od 5 °C do 30 °C. Podmínky jsou podobné P. graminis, ale s nižším teplotním prahem 5 °C.
Pokud se prokáže infekce 100%, mělo by se zvážit kurativní měření ochrany rostlin. Pokud je riziko na úrovni 80% a předpověď počasí předpovídá další období vlhnutí listů, mohly by být provedeny ochranné aplikace na listy.
Vlhkost listů pro kumulované průměrné hodinové teploty pro 90 °C:
- (pokud T <= 22,5 °C, pak ∑(Th) jinak ∑ (22,5-(Th-22,5))
- 5°C < Temp. < 30°C
Model infekce Puccinia graminis
K infekci dochází po několika hodinách navlhčení listů za optimálních teplotních podmínek. Model předpokládá, že infekce potřebuje kumulativní hodinovou teplotu 80 °C vlhkosti listů v rozmezí teplot vzduchu od 10 °C do 35 °C. Upřednostňuje o něco vyšší teplotu než P. triticina a po infekci musí následovat sluneční světlo.
Pokud je prokázána infekce 100%, podmínky jsou pro houbu příznivé, a proto je třeba zvážit opatření na ochranu rostlin.
Vlhkost listů pro kumulované průměrné hodinové teploty 80 °C následované světelným obdobím (150 W/m²) pro kumulované průměrné hodinové teploty 30 °C:
- (pokud T <= 24 °C, pak ∑(Th) jinak ∑ (Th-24))
- 10°C < Temp. < 35°C
Model infekce Puccinia striiformis
Puccinia striiformis je rez pšeničná chladného podnebí, která má optimální teplotu již od 15 °C. K infekci dochází po několika hodinách navlhčení listů při optimálních teplotních podmínkách. Model předpokládá, že infekce potřebuje kumulativní hodinovou teplotu 80 °C vlhkosti listů v rozmezí teplot vzduchu od 5 °C do 20 °C. V obdobích s nízkou intenzitou světla k infekcím nedochází.
Než graf dosáhne hodnoty 100%, je možné provést ochrannou aplikaci, zatímco po jejím dosažení je třeba zvážit léčebné ochranné strategie (systémové).
Vlhkost listů a světlo pro kumulované hodinové průměrné teploty pro 80 °C:
- (pokud T <= 15 °C, pak ∑(Th) jinak ∑ (Th-15))
- 5°C < Temp. < 20°C
Literatura
- Bolton, M. D., Kolmer, J. A., & Garvin, D. F. (2008). Rez listová pšenice způsobená houbou Puccinia triticina. Molekulární patologie rostlin, 9(5), 563-575.
- Chen, X. M. (2005). Epidemiologie a kontrola rzi pruhované [Puccinia striiformis f. sp. tritici] na pšenici. Canadian journal of plant pathology, 27(3), 314-337.
- Figueroa, M., Hammond-Kosack, K. E., & Solomon, P. S. (2018). A review of wheat diseases-a field perspective (Přehled chorob pšenice - pohled z terénu). Molekulární patologie rostlin, 19(6), 1523-1536.
- Průvodce, A. Choroby pšenice způsobené rzí listovou, stonkovou a pruhovanou. Leonard, K. J., & Szabo, L. J. (2005). Stébelná rez drobných obilovin a trav způsobená houbou Puccinia graminis. Molekulární patologie rostlin, 6(2), 99-111.
- Kolmer, J. (2013). Leaf rust of wheat: pathogen biology, variation and host resistance (Rezistence pšenice k listům: biologie patogenu, variabilita a odolnost hostitele). Lesy, 4(1), 70-84.
- Singh, R. P., Huerta-Espino, J., Roelfs, A. P., & Curtis, B. C. (2002). Pšenice rezaví. Růst, 2(25), 35.