

Ранній фітофтороз картоплі та томатів
Рендалл К. Роу, Саллі А. Міллер, Річард М. Рідел, Дорадча служба Університету штату Огайо
Фітофтороз є дуже поширеним захворюванням як картоплі, так і томатів. Вона викликає плямистість листя і бульб у картоплі, а у томатів - плямистість листя, плодову гниль і ураження стебла. Захворювання може виникати в широкому діапазоні кліматичних умов і може бути дуже руйнівним, якщо його не контролювати, часто призводячи до повної дефоліації рослин. Всупереч назві, вона рідко розвивається на ранніх стадіях, але зазвичай з'являється на зрілому листі.
Симптоми
На листках обох культур перші симптоми зазвичай з'являються на старих листках і являють собою невеликі, неправильної форми, від темно-коричневого до чорного кольору, мертві плями розміром від крапки до 1/2 дюйма в діаметрі. Коли плями збільшуються, можуть утворюватися концентричні кільця внаслідок нерегулярного росту організму в тканині листка. Це надає ураженню характерного вигляду "плями-мішені" або "яблучне око". Навколо кожної плями часто є вузький жовтий ореол, а місця ураження, як правило, облямовані жилками. Коли плям багато, вони можуть зростатися, що призводить до пожовтіння та відмирання заражених листків. Зазвичай першими заражаються найстаріші листки, які засихають і опадають з рослини в міру того, як хвороба прогресує вгору по головному стеблу.
На томатах стеблові інфекції можуть виникати в будь-якому віці, що призводить до появи невеликих темних, злегка заглиблених ділянок, які збільшуються, утворюючи круглі або витягнуті плями зі світлішими центрами. На стеблах часто утворюються концентричні плями, схожі на ті, що на листках. Якщо для пересадки томатів використовується заражене насіння, розсада може засохнути незабаром після появи сходів. Коли на стеблах пересаджених рослин або розсади розвиваються великі ураження на рівні ґрунту, рослини можуть бути опоясані - стан, відомий як "шийна гниль". Такі рослини можуть загинути під час висадки в поле або, якщо стебла ослаблені, зламатися на початку сезону. Деякі рослини можуть вижити зі зменшеною кореневою системою, якщо частини стебел над раком розвинуть коріння в місцях контакту з ґрунтом. Такі рослини, однак, зазвичай дають мало плодів або взагалі не дають плодів. Ураження стебла набагато рідше зустрічається на картоплі і є руйнівним для неї.
Коли рання гниль вражає томати на стадії цвітіння, може спостерігатися опадання цвіту і плямистість плодових стебел, а також втрата молодих плодів. На старих плодах фітофтороз викликає темні, шкірясті вдавлені плями, зазвичай в місці прикріплення плодоніжки. Ці плями можуть збільшуватися і охоплювати всю верхню частину плоду, часто показуючи концентричні позначки, подібні до тих, що є на листі. Уражені ділянки можуть бути вкриті оксамитовою чорною масою спор. Плоди також можуть бути інфіковані на зеленій або стиглій стадії через ростові тріщини та інші рани. Заражені плоди часто опадають, не досягнувши зрілості.
На бульбах картоплі рання фітофтороза призводить до поверхневих уражень, які виглядають трохи темнішими, ніж прилегла здорова шкіра. Ураження, як правило, злегка заглиблені, круглі або неправильної форми і варіюються в розмірах до 3/4 дюйма в діаметрі. Зазвичай між здоровою та ураженою тканиною є чітко окреслена, а іноді трохи піднята межа. Зсередини тканина має коричневу або чорну коркову суху гниль, зазвичай глибиною не більше 1/4 - 3/8 дюйма. На старих ураженнях можуть утворюватися глибокі тріщини. Бульбова інфекція в умовах Огайо зустрічається рідко.
Збудник
Ранній фітофтороз викликається грибком Alternaria solani, який виживає в інфікованих тканинах листя або стебла на ґрунті або в ґрунті. Цей грибок повсюдно присутній на полях, де вирощують ці культури. Він також може переноситися на насінні томатів та бульбах картоплі. Спори утворюються на заражених рослинних рештках на поверхні ґрунту або на активних уражених ділянках у досить широкому діапазоні температур, особливо в умовах чергування вологих і сухих умов. Вони легко переносяться повітряними потоками, вітром, дощовими бризками та поливною водою. Зараження сприйнятливих тканин листя або стебла відбувається в теплу, вологу погоду з рясними росами або дощами. Ранній фітофтороз може розвиватися досить швидко в середині та наприкінці сезону і є більш серйозним, коли рослини перебувають у стресовому стані через погане живлення, посуху або інших шкідників. Зараження бульб картоплі відбувається через природні отвори на шкірці або через пошкодження. Бульби можуть контактувати зі спорами під час збирання врожаю, і ураження може продовжувати розвиватися під час зберігання.
TOMCAST (Джим Ясінські, координатор TOMCAST в Огайо, Індіані та Мічигані)
Передумови
TOMCAST (TOMato disease foreCASTing) - це комп'ютерна модель, заснована на польових даних, яка намагається передбачити розвиток грибкових захворювань, а саме раннього фітофторозу, септоріозу листя та антракнозу на томатах. Реєстратори даних, розміщені на полях, реєструють погодинні дані про вологість і температуру листя. Ці дані аналізуються протягом 24 годин і можуть призвести до формування Значення тяжкості захворювання (DSV); по суті, це приріст розвитку хвороби. У міру накопичення DSV, тиск хвороб продовжує наростати на культуру. Коли кількість накопичених DSV перевищує інтервал між обприскуваннями, рекомендується застосування фунгіциду, щоб зменшити тиск хвороби.
TOMCAST
Час застосування фунгіцидів проти раннього фітофторозу, септоріозу листя та антракнозу
Система прогнозування хвороб на основі погодних умов під назвою TOMCAST, розроблена доктором Роном Пітбладо в Ріджтаунському коледжі сільськогосподарських технологій в Онтаріо, Канада, може бути використана для визначення часу застосування фунгіцидів проти трьох грибкових захворювань: раннього фітофторозу (викликається Alternaria solani), септоріозної плямистості листя (викликається Septoria lycopersici) та антракнозу плодових (викликається Colletotrichum coccodes). Якщо фітофтороз присутній у вашому або сусідніх округах, або існують умови для переміщення спор на вашу територію, використовуйте систему прогнозування фітофторозу Simcast, щоб визначити час застосування фунгіцидів.
ВАЖЛИВЕ ЗАСТЕРЕЖЕННЯ:
ТОМКАСТ не є корисним на фермах, які мають історію бактеріальних захворювань. Якщо у вас часто виникають проблеми з бактеріальною плямистістю, плямистістю або раком на томатах, вам не слід використовувати ТОМКАСТ, оскільки рекомендовані інтервали між обприскуваннями будуть недостатніми для контролю бактеріальних захворювань, якщо ви змішуєте мідь з фунгіцидами в баках. Знайдіть інформацію про зменшення бактеріальних захворювань тут: http://extension.psu.edu/plants/vegetable-fruit/news/2015/farming-like-you-expect-bacterial-diseases
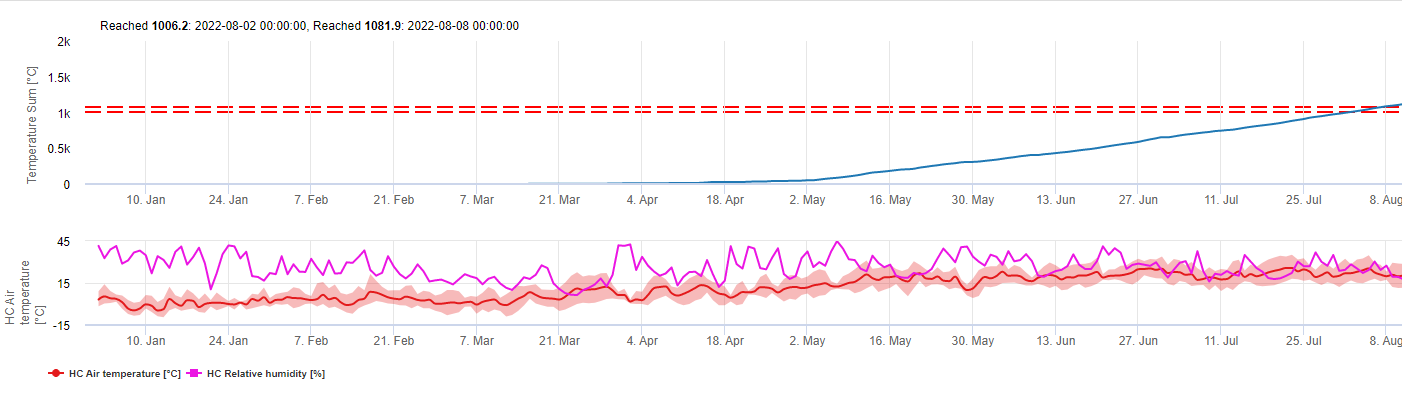
TOMCAST використовує дані про вологість і температуру листя для розрахунку показників тяжкості хвороби (DSV), як показано в Таблиці 1.
TOMCAST походить від оригінальної моделі F.A.S.T. (Forecasting Alternaria solani on Tomatoes), розробленої докторами Медденом, Пенніпекером і Макнабом в Університеті штату Пенсильванія (PSU). Модель PSU F.A.S.T. була в подальшому модифікована доктором Пітбладо в коледжі Ріджтаун в Онтаріо в те, що ми зараз визнаємо як модель TOMCAST, яка використовується в Університеті штату Огайо.
DSV Значення тяжкості хвороби (DSV) - це одиниця виміру, що відповідає певному інкременту розвитку хвороби (раннього фітофторозу). Іншими словами, DSV - це числове представлення того, наскільки швидко або повільно хвороба (ранній фітофтороз) накопичується на томатному полі. DSV визначається двома факторами: вологістю листя і температурою в години "мокрого листя". Зі збільшенням кількості годин з мокрим листям і температури DSV накопичується швидше. Нижче наведено діаграму "Тяжкість хвороби".
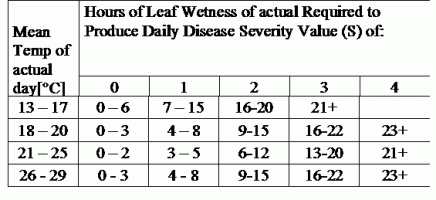
І навпаки, за меншої кількості годин зволоження листя та нижчої температури DSV накопичуються повільніше, якщо взагалі накопичуються. Коли загальна кількість накопичених DSV перевищує встановлену межу, яка називається інтервалом або порогом обприскування, рекомендується обприскування фунгіцидом, щоб захистити листя і плоди від розвитку хвороб.

Інтервал між обприскуваннями (який визначає, коли слід обприскувати) може коливатися в межах 15-20 DSV. Точне значення DSV, яке повинен використовувати виробник, зазвичай надається переробником і залежить від якості плодів і кінцевого використання томатів. Дотримання інтервалу обприскування 15 DSV є консервативним використанням системи TOMCAST, тобто ви будете обприскувати частіше, ніж фермер, який використовує інтервал обприскування 19 DSV з системою TOMCAST. Компроміс полягає в кількості обприскувань протягом сезону і потенційній різниці в якості плодів.
ВИКОРИСТАННЯ TOMCAST
Томати, вирощені в радіусі 10 миль від станції спостереження, можуть скористатися функцією управління хворобами TOMCAST для прогнозування раннього фітофторозу, септоріозу та антракнозу. Якщо ви вирішите спробувати TOMCAST в цьому сезоні, будь ласка, майте на увазі три дуже важливі концепції.
Перше: Якщо ви вперше використовуєте систему, рекомендується внести в програму лише частину ваших посівних площ, щоб побачити, наскільки вона відповідає вашим стандартам якості та стилю роботи.
Друге: використовуйте TOMCAST як орієнтир для вибору часу внесення фунгіцидів, розуміючи, що в деякі сезони ви можете внести більше продукту, ніж може вимагати програма з встановленим графіком.
Третє: Чим далі томатне поле знаходиться від місця збору даних, тим більша ймовірність викривлення в накопиченні DSV, тобто звітне значення може бути на кілька DSV вищим або нижчим від того, що спостерігається на місці розташування поля. Це слід враховувати, коли застосування фунгіцидів, ймовірно, відбудеться через кілька днів. Прослухайте звіти про DSV сусідніх станцій і проведіть тріангуляцію до свого місця розташування - це найкращий спосіб приблизно оцінити накопичення DSV.
ПЕРШЕ ОБПРИСКУВАННЯ З ВИКОРИСТАННЯМ TOMCAST
Протягом багатьох років точилися дискусії щодо застосування першого обприскування при використанні ТОМКАСТ. Правило, викладене в Посібнику з овочівництва 1997 року, зосереджується на даті посадки.
Рослини томатів, висаджені в поле до 20 травня, слід обприскувати, коли DSV для цієї зони перевищує 25 або коли настає безпечна дата - 15 червня. Безпечна дата використовується тільки в тому випадку, якщо ви не проводили обприскування з 20 травня, і є засобом для усунення початкового зародка хвороби. Після першого обприскування ці томати обробляють, коли перевищується обраний інтервал обприскування (діапазон 15-20 DSV).
Томати, висаджені після 20 травня, обробляють, коли вони перевищують обраний інтервал обприскування (діапазон 15-20 DSV) або коли вони не були оброблені до безпечної дати 15 червня. Тому дуже важливо порівняти дату посадки томатів з датою початку звітності по DSV на цій ділянці, щоб керуватися нею в процесі прийняття рішення про обприскування.
Існує дві основні септоріозні хвороби пшениці. Це септоріозна плямистість, спричинена грибком Septoria tritici Septoria tritici (телеоморф: Mycophaerella graminicola), та Септорія вузлувата (Septoria nodorum) плямистості, спричиненої грибком Septoria nodorum (телеоморф: Leptosphaeria nodorum). Обидві хвороби спричиняють серйозні втрати врожаю, які коливаються від 31 до 53% (Eyal, 1981; Babadoost and Herbert, 1984; Polley and Thomas, 1991). У всьому світі уражено понад 50 млн га пшениці, яка переважно вирощується в районах з великою кількістю опадів. Протягом останніх 25 років ці хвороби зростали і стали основним обмежувальним фактором для виробництва пшениці в певних регіонах. Під час сильних епідемій зерна сприйнятливих сортів пшениці стають щуплими і не придатними для помелу. Епідемії септоріозної та нодорозної плямистостей пшениці пов'язані зі сприятливими погодними умовами (часті дощі та помірні температури), специфічними культуральними практиками, доступністю посівного матеріалу та наявністю сприйнятливих сортів пшениці (Eyal et al., 1987).
Біологія септоріозу
За Еріком де Вольфом, пляма Septoria Tritici, Університет штату Канзас, квітень 2008 року Septoria tritici плямистість, відома як крапчаста плямистість листя, спричинена грибком Septoria tritici. Вона поширена в усіх регіонах світу, де вирощують пшеницю, і є серйозною проблемою в багатьох регіонах. Найбільшої шкоди септоріозна плямистість завдає, коли хвороба вражає верхні листки і качани сприйнятливих сортів наприкінці сезону.
Симптоми
Septoria tritici Симптоми плямистості вперше з'являються восени. Початковими симптомами є невеликі жовті плями на листі. З віком ці ураження часто набувають світло-коричневого відтінку, а плодові тіла гриба можна помітити на остюках, вбудовані в уражені ділянки. Ураження мають неправильну форму і варіюються від еліптичних до довгих і вузьких (рис. 1). Ураження містять маленькі, круглі, чорні цятки, які є плодовими тілами гриба. Чорні плодові тіла схожі на зерна чорного перцю і зазвичай їх можна побачити без допомоги лупи. Хвороба починається з нижніх листків і поступово переходить на прапорцевий листок. Листові піхви також схильні до ураження. У вологі роки крапчаста плямистість листя може переходити на качани і викликати коричневі ураження на колоссях і остюках, відомі як плямистість колосся. З віком ці ураження часто набувають світло-коричневого кольору, а плодові тіла гриба часто можна побачити вбудованими в ураження на остюках.
Фаза плямистості може спричинити значні втрати врожаю, але взаємозв'язок між тяжкістю хвороби та втратами врожаю недостатньо вивчений. Septoria tritici плямистість можна сплутати з іншими хворобами листя, які мають дуже схожі симптоми: плямистістю та Stagonspora nodorum наприклад, плямистості. Зазвичай рослини уражуються більш ніж однією з цих хвороб листя, і для точного діагностування найбільш поширених хвороб може знадобитися лабораторне обстеження. Лабораторне дослідження майже завжди потрібне, щоб визначити причину появи плямистостей. Знання виду не є важливим для прийняття рішення щодо обприскування, оскільки всі три хвороби однаково реагують на фунгіциди. Однак знання того, які хвороби найбільш поширені, є важливою частиною селекції сортів, оскільки різні гени контролюють стійкість до хвороб.
Найнадійніший спосіб відрізнити плямистість Septoria tritici від інших захворювань - це наявність чорних плодових тіл гриба. Гриб, що спричиняє засмагу, не утворює такого типу репродуктивних структур. Однак у вологих умовах гриб, що викликає плямистість Stagonospora nodorum, утворює світло-коричневі плодові тіла. Крім різниці в кольорі, ці структури також менші, ніж ті, що утворює Septoria tritici.
Життєвий цикл
Septoria tritici виживає влітку на залишках попереднього врожаю пшениці та ініціює інфікування восени. Існують певні докази того, що гриб здатний виживати в асоціації з іншими рослинами-господарями та насінням пшениці. Ці джерела гриба, ймовірно, є найбільш важливими за відсутності пшеничних решток. Незалежно від сівозміни або практики управління пожнивними рештками, зазвичай є достатньо посівного матеріалу, щоб ініціювати осінню інфекцію. Septoria tritici плямистості сприяє прохолодна волога погода. Оптимальний температурний діапазон - від 16 до 21 °C, проте інфікування може відбуватися і взимку за температури до 5 °C. Для зараження потрібно, щоб листя було вологим щонайменше 6 годин, а для максимального зараження - до 48 годин. Після зараження грибу потрібно від 21 до 28 днів, щоб розвинути характерні чорні плодові тіла і виробити нове покоління спор. Спори, що утворюються в цих плодових тілах, виділяються у вигляді липкої маси і потребують дощу, щоб потрапити на верхні листки і качани.
Зараження через Septoria tritici
Пікнідіоспори S. tritici проростають у вільній воді з обох кінців спори або з інтеркалярних клітин (Weber, 1922). Проростання спор починається приблизно через 12 годин після контакту з листком. Зародкові трубки ростуть хаотично по поверхні листка. Вебер (1922) спостерігав лише пряме проникнення між клітинами епідермісу, але інші дослідники дійшли висновку, що проникнення як через відкриті, так і через закриті продихи є основним способом проникнення в хазяїна (Benedict, 1971; Cohen and Eyal, 1993; Hilu and Bever, 1957). Кема та ін. (1996) спостерігали лише продихове проникнення. Гіфи, що проростають через продихи, звужуються до діаметру близько 1 мкм, а потім стають ширшими після досягнення підпродихової порожнини.
Гіфи ростуть паралельно поверхні листка під клітинами епідермісу, потім через мезофіл до клітин нижнього епідермісу, але не в епідерміс. Гаусторії не утворюються, а ріст гіф обмежується клітинами склеренхіми навколо судинних пучків, за винятком випадків, коли гіфи дуже щільні. Судинні пучки не інвазуються. Гіфи ростуть міжклітинно вздовж клітинних стінок через мезофіл, розгалужуючись на перегородці або в середині клітини. Макроскопічні симптоми не з'являються протягом приблизно 9 днів, за винятком випадкових мертвих клітин, але клітини мезофілу швидко гинуть через 11 днів. Пікніди розвиваються в субтоматичних камерах. Гіфи рідко проростають у клітини хазяїна (Hilu and Bever, 1957; Kema et al, 1996; Weber, 1922).
Успішне зараження відбувається лише після щонайменше 20 годин високої вологості. Лише кілька коричневих плям розвиваються, якщо листя залишається вологим протягом 5-10 годин після осадження спор (Holmes and Colhoun, 1974) або до 24 годин (Kema et al., 1996). Відносини хазяїн-паразит є однаковими як на стійких, так і на сприйнятливих пшеницях. Проростання спор на поверхні листя однакове незалежно від сприйнятливості. Кількість успішних проникнень приблизно однакова, але ріст гіф відбувається швидше на сприйнятливих сортах, що призводить до більшого ураження. Гіфи поширюються 44 Сесія 2 - Б.М. Кунфер за межі некротичних ділянок у всіх сортів. Токсин може відігравати певну роль у патогенезі (Cohen and Eyal, 1993; Hilu and Bever, 1957). На противагу цьому, колонізація була значно знижена на стійкій лінії (Kema et al., 1996).
Стагоноспори (септоріоз) та збудники септоріозу зернових: Інфекційний процес
Б.М. Канфер, кафедра патології рослин, Університет Джорджії, Гріффін, штат Джорджія
Процес зараження найбільш інтенсивно вивчався для Stagonospora (Septoria) nodorum та Septoria tritici. Одне поглиблене дослідження на тему Septoria passerinii є доступною. Майже вся інформація стосується інфікування пікнідоспорами. Однак процес інфікування іншими споровими формами досить схожий. Представлена інформація здебільшого стосується інфікування листків за оптимальних умов. Деякі дослідження були проведені з інтактними проростками, тоді як інші - з відокремленими листками. Інфікування колеоптиля та проростка пшениці S. nodorum був детально описаний Бейкером (1971) і переглянутий Кунфером (1983). Хоча точних порівнянь не проводилося, схоже, що інфекційний процес має багато спільного в кожній системі хазяїн-паразит і є типовим для багатьох некротрофних патогенів. Інформація про фактори, що впливають на розвиток симптомів і вираженість хвороби, не включена, але була розглянута іншими авторами (Eyal et al., 1987; King et al., 1983; Shipton et al., 1971). Включено короткий опис факторів, що впливають на тривалість життя спор на поверхні листка.
Роль циркуляції та виживання спор на поверхні листка Найбільш детальна інформація про функцію кільця, що оточує пікнідоспори, які виділяються з пікнідія, міститься у S. nodorum. Циррус - це гель, що складається з білкових і сахаридних сполук. Його склад і функції подібні до складу та функцій інших грибів Sphaeropsidales (Fournet, 1969; Fournet et al., 1970; Griffiths and Peverett, 1980). Основними функціями компонентів цирруса є захист пікнідіоспор від десикації та запобігання передчасному проростанню.
Циррус захищає пікнідоспори так, що деякі з них залишаються життєздатними щонайменше 28 днів (Fournet, 1969). Коли циррус розводили водою, якщо концентрація розчину цирруса була >20%, проростало менше 10% пікнідіоспор. При меншій концентрації компоненти забезпечують поживні речовини, які стимулюють проростання спор і подовження зародкових трубок. Довжина зародкових трубок збільшувалася до концентрації цирруса 15%, а потім помірно зменшувалася за вищих концентрацій (Harrower, 1976). Бреннан та ін. (Brennan et al., 1986) повідомили про більшу схожість у розведеній рідині цирруса. Компоненти вихору знижували схожість при відносній вологості 10-60%. Після того, як спори дисперговані, стимулюючий вплив циррусної рідини, ймовірно, є незначним (Гріффітс і Певеретт, 1980).
При відносній вологості 35-45% спори S. tritici в циррі залишалися життєздатними щонайменше 60 днів (Gough and Lee, 1985). Компоненти цирруса можуть діяти як інгібітори проростання спор, або ж високий осмотичний потенціал цирруса може перешкоджати проростанню. Пікнідіоспори S. nodorum не виживали протягом 24 годин при відносній вологості вище 80% при 20 C. Спори виживали два тижні або більше при відносній вологості <10% (Griffiths and Peverett, 1980). Коли рідина цирруса S. nodorum розбавляли водою, близько двох третин пікнідоспор втрачали життєздатність протягом 8 годин, а через 30 годин при денному світлі проростала лише 5%. Коли спори зберігалися в темряві, 40% залишалися життєздатними через 30 годин (Brennan et al., 1986).
Сухі конідії S. nodorumу затінку та під прямими сонячними променями виживали на відкритому повітрі щонайменше 56 годин (Fernandes and Hendrix, 1986a). Проростання пікнідоспор S. nodorum пригнічувалося безперервним УФ-В (280-320 нм), тоді як проростання S. tritici не було. Подовження зародкових трубок під впливом безперервного ультрафіолетового випромінювання пригнічувалося для обох грибів порівняно з темрявою (Rasanayagam et al., 1995).
Зараження Septoria nodorum
Процес проникнення в організм хазяїна і розвиток S. nodorum в листі було детально вивчено кількома дослідниками (Baker and Smith, 1978, Bird and Ride 1981, Karjalainen and Lounatmaa, 1986; Keon and Hargreaves, 1984; Straley, 1979; Weber, 1922). Пікнідіоспори, як правило, розташовуються в заглибленнях між двома епідермальними клітинами, і багато спроб проникнення в листки починаються саме там. Спори проростають на поверхні листка у відповідь на вільну вологу (Fernandes and Hendrix, 1986b). Вони починають проростати через 2-3 години після осадження, а через 8 годин проростання може досягати 90%. Проникнення в листя починається приблизно через 10 годин після осадження спор (Bird and Ride, 1981; Brönnimann et al., 1972; Holmes and Colhoun, 1974).
На початку проростання зародкова трубка оточена аморфним матеріалом, який прикріплюється до листка. Зародкові трубки, що ростуть з обох кінців спори та з інтеркалярних клітин, мають тенденцію рости вздовж заглиблень між клітинами і часто орієнтовані вздовж довгої осі листка (O'Reilly and Downes, 1986). Гіфи зі спор, що не знаходяться в заглибленнях, ростуть хаотично з рідкісним розгалуженням (Straley, 1979). Утворюється аппресорій з інфекційним кілочком, який проникає крізь кутикулу і периклінальну стінку епідермальних клітин безпосередньо в клітинний просвіт, що призводить до швидкої загибелі клітини.
У багатьох випадках проникнення спочатку відбувається субкутикулярно або бічний ріст гіфи всередині клітинної стінки, перш ніж вона проростає в цитоплазму (Bird and Ride, 1981; O'Reilly and Downes, 1986). Проникнення як через відкриті, так і через закриті продихи також відбувається і може бути швидшим, ніж пряме проникнення (Harrower, 1976; Jenkins, 1978; O'Reilly and Downes, 1986; Straley, 1979). Зародкові трубки розгалужуються на продихах і з'єднаннях клітин епідермісу. Проникнення зародкової трубки в продихи може відбуватися без утворення продиху. Іноді проникнення відбувається через трихоми (Straley, 1979). Очевидно, більшість спроб проникнення є невдалими, оскільки в клітинах на місці спроби проникнення утворюються щільні сосочки (Karjalainen and Lounatmaa, 1986; Bird and Ride, 1981).
Після проникнення клітини епідермісу швидко відмирають і здерев'яніють, а гіфи вростають у мезофіл. Клітини мезофілу деформуються, і здерев'янілий матеріал відкладається поза деякими клітинами, які потім руйнуються. Здерев'яніння відбувається до того, як гіфи досягнуть клітини. Процес однаковий у всіх сортів, але у стійких сортів розвивається повільніше. Гіфи ростуть міжклітинно між клітинами епідермісу, а потім у мезофіл. Після проникнення в мезофіл руйнування хлоропластів починається через 6-9 днів (Karjalainen and Lounatmaa, 1986).
Однак швидкість фотосинтезу починає знижуватися вже через добу після зараження і до появи симптомів (Krupinsky et al, 1973). Склеренхімна тканина навколо судинних пучків запобігає інфікуванню судинної тканини. Судинні пучки блокують поширення гіфів через мезофіл, за винятком випадків, коли тканина склеренхіми молода і не повністю сформована (Baker and Smith, 1978).
Stagonospora nodorum виділяє широкий спектр ферментів, що руйнують клітинну стінку, включаючи амілазу, пектинметилестеразу, полігалактуронази, ксиланази та целюлазу in vitro та під час зараження листя пшениці (Baker, 1969; Lehtinen, 1993; Magro, 1984). Інформація, пов'язана з деградацією клітинної стінки ферментами, узгоджується з гістологічними спостереженнями. Ці ферменти можуть діяти разом з токсинами. Чутливість до ферментів може бути пов'язана зі стійкістю та швидкістю колонізації грибів (Magro, 1984). Як і багато некротрофів, збудники септоріозу та стагоноспори продукують фітотоксичні сполуки in vitro. Погіршення стану і загибель клітин до початку росту гіфів у тканину мезофілу (Bird and Ride, 1981) узгоджується з продукуванням токсинів. Однак, остаточна роль токсинів в інфекційному процесі та їх зв'язок зі стійкістю хазяїна не встановлені (Bethenod et al, 1982; Bousquet et al, 1980; Essad and Bousquet, 1981; King et al, 1983). Відмінності в діапазоні господарів між адаптованими до пшениці та ячменю штамами S. nodorum може бути пов'язана з виробленням токсинів (Bousquet і Kollmann, 1998). Ініціація проростання спор і відсоток пророслих спор не залежать від сприйнятливості хазяїна (Bird and Ride, 1981; Morgan 1974; Straley, 1979; Straley and Scharen, 1979; Baker and Smith, 1978).
Bird і Ride (1981) повідомили, що розширення зародкових трубок на поверхні листя відбувалося повільніше на стійких, ніж на сприйнятливих сортах. Цей механізм, що проявляється щонайменше через 48 годин після осадження спор, вказує на стійкість до проникнення до подовження зародкових трубочок. На стійких сортах було менше успішних проникнень, і проникнення відбувалося повільніше на стійких сортах (Baker and Smith, 1978; Bird and Ride, 1981). Було запропоновано лігніфікацію для обмеження інфекції як у стійких, так і у сприйнятливих сортів, але інші фактори сповільнювали розвиток гриба на стійких лініях. У сприйнятливих лініях гіфи, що швидше ростуть, можуть уникати лігніфікації клітин хазяїна. Через чотири дні після інокуляції ячменю ізолятом пшеничного біотипу гриба S. nodorumгіфи проростали крізь кутикулу, а іноді і в зовнішні целюлозні шари клітинних стінок епідермісу. Товсті сосочки відкладалися під проникними гіфами, і клітини не проникали (Keon and Hargreaves, 1984).
Зараження Septoria passerinii: Грін і Діксон (Green and Dickson, 1957) представили детальний опис процесу зараження S. passerinii на ячмені. Процес зараження схожий на S. tritici. Як і для S. tritici, час, необхідний для проникнення в листя, значно довший, ніж для S. nodorum. Зародкові трубки розгалужуються і ростуть по поверхні листка хаотично, але іноді вздовж заглиблень між клітинами епідермісу. Проникнення в листки відбувається майже виключно через продихи. Проростаючі гіфи набухають, і якщо проникнення не вдається, гіфи продовжують витягуватися. Через 48 годин після осадження спор проникнення не відбувається. Через 72 години зародкові трубки потовщуються над продихами, ростуть між охоронними клітинами, на поверхнях додаткових клітин і проникають у підпродихові порожнини. Пряме проникнення між клітинами епідермісу спостерігається дуже рідко.
Проростання спор і проникнення в рослину є однаковими на стійких і сприйнятливих сортах. На стійких сортах спостерігається значно менше поширення гіф всередині листя, а сосочки спостерігаються на багатьох, але не на всіх клітинних стінках. Гіфи ростуть під епідермою від одного продиху до іншого, але не проникають між клітинами епідерми. Мезофіл колонізується, але гаусторії не утворюються. Після того, як клітини мезофілу некротизуються, клітини епідермісу руйнуються. Розвиток міцелію в листі рідкісний і зазвичай блокується судинними пучками. У молодих листках, якщо судинна оболонка менш розвинена, гіфи проходять між пучком і епідермісом. Пікніди утворюються в субтоматних порожнинах, переважно на верхній поверхні листка (Green and Dickson, 1957).
Фактори, що впливають на довговічність спор на листковій поверхні Серед збудників стагоноспорозу та септоріозу зернових культур достовірна інформація про інфекційний процес була отримана лише для S. nodorum, S. triticiі S. passerinii. Як і багато інших некротрофних патогенів, жодна з груп збудників не викликає гіперчутливої реакції. Істотна відмінність в інфекційному процесі між збудниками септоріозу та стагоноспори полягає в тому, що проростання і проникнення спор відбувається набагато швидше для збудників септоріозу S. nodorum ніж для S. tritici і S. passerinii. Це має значний вплив на епідеміологію захворювань.
Збудники септоріозу проникають у рослину переважно через продихи, тоді як S. nodorum проникає як безпосередньо, так і через продихи. S. nodorum швидко проникає і вбиває клітини епідермісу, але S. tritici та S. passerinii не вбивають клітини епідермісу, поки гіфи не проростуть через мезофіл листка і не почнеться швидкий некроз. Гістологічні дослідження росту гриба після проникнення в організм хазяїна збігаються з даними епідеміологічних досліджень резистентності хазяїна. Резистентність сповільнює швидкість колонізації хазяїна, але не має помітного впливу на процес розвитку ураження.
Механізми, що контролюють реакцію хазяїна, пов'язані з ферментами, токсинами або іншими метаболітами, що вивільняються патогенами під час інфікування, все ще залишаються незрозумілими. Існує мало інформації про інфікування аскоспорами. Процес зараження, ймовірно, дуже схожий на процес зараження пікнідоспорами. Аскоспори Phaeosphaeria nodorum проростають в широкому діапазоні температур, а їхні зародкові трубки проникають безпосередньо в листок. Однак, за даними Rapilly та ін. (1973), аскоспори, на відміну від пікнідоспор, не проростають у вільній воді.
Модель інфекції, спричиненої септоріозом
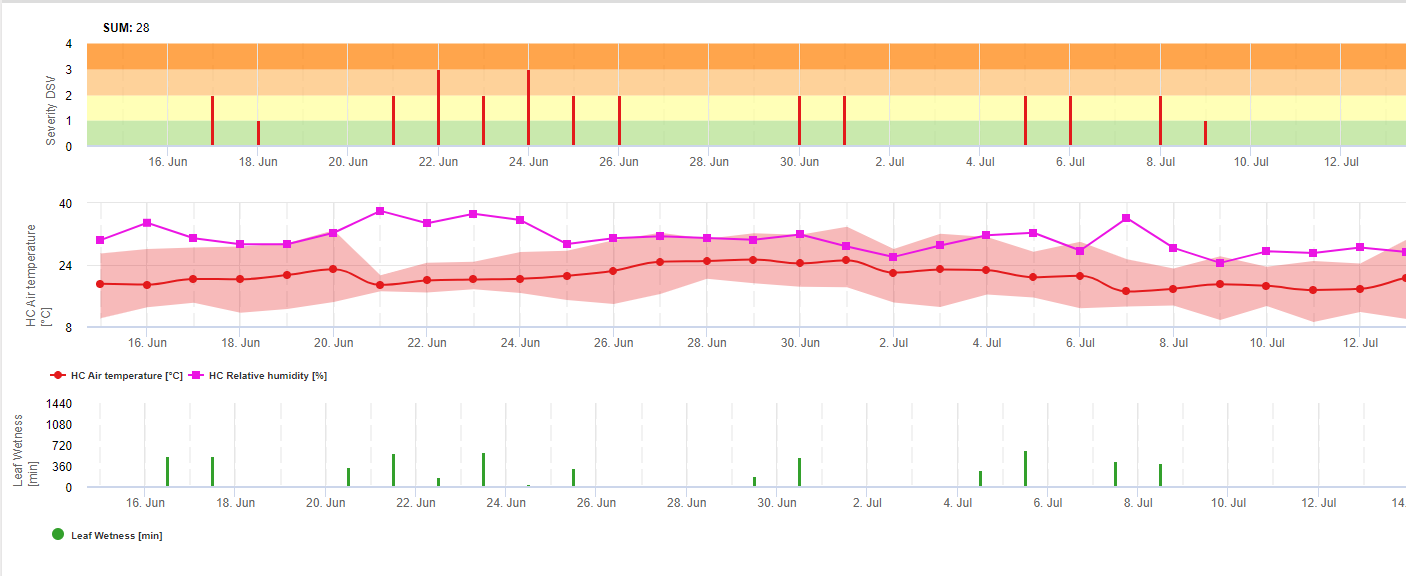
Зараження септоріозом можливе при низьких температурах, тоді як температура нижче 7°C може не призвести до інфікування протягом 2 днів. Оптимальна температура для розвитку хвороби досягається в діапазоні від 16 до 21°C. Зараження можливе в період високої відносної вологості повітря або вологості листя протягом 14 годин або довше. Щоб відповідати цим умовам, ми вирішили розділити моделі на слабкі, помірні та сильні інфекції. Слабкі інфекції можуть бути задані, якщо патоген може інфікувати тканину хазяїна. Це означає, що слабкі інфекції можуть мати місце, якщо температури мінімальні, а періоди зволоження листя мають критичну тривалість. Помірне інфікування відбувається за умов, коли більшість випробувань на інфекційність призводять до задовільних результатів, а сильне інфікування відбувається за умов, коли патоген має оптимальні умови для інфікування.

Починається інфікування після дощу 0,5 мм. Ми вирішили не використовувати модель утворення пікнід. Умовою, необхідною для утворення пікнід, вважається період з відносною вологістю повітря вищою за 85%. Очікується, що час життя пікнід становить 24 години. У всіх кліматичних зонах, де Septoria tritici має можливість інфікувати, ми знайдемо 2 години, що відповідають цим умовам, майже кожного дня на сході сонця.
Оцінка тяжкості інфекції: Для того, щоб мати змогу оцінити інфекційний тиск Septoria tritici між стадіями 10 (колеоптиль першого листкового жолоба) і 32 (другий вузол принаймні на 2 см вище вузла 1), а також між 32 і 51 (початок колосіння), ми повинні оцінити ступінь тяжкості інфекції, виходячи з кліматичних умов. Ця оцінка проводиться за шкалою від 1 до 5. Ступінь тяжкості 1 ставиться, якщо умови для слабкої інфекції виконані, а дощ випав менше 5 мм, в іншому випадку відповідний показник тяжкості буде дорівнювати 2. Ступінь важкості 3 присвоюється, якщо виконані умови для помірного зараження і випало менше 5 мм опадів. Якщо під час помірного зараження випало більше 5 мм опадів, а під час сильного зараження - менше 5 мм опадів, то ступінь вираженості інфекції дорівнює 4.
Сильна інфекція з кількістю опадів понад 5 мм відповідає значенню ступеня тяжкості 5.
Septoria tritici оцінка тиску хвороби: Клімат - це лише один з факторів, що впливає на розвиток хвороб на полях. Іншими двома факторами є історія поля та сприйнятливість вирощуваного сорту. Якщо ми можемо накопичити значення тяжкості хвороби від стадії 10 до стадії 32 до значення 4, ми можемо очікувати слабкого тиску хвороби з боку клімату. Якщо це значення досягає 6, ми можемо очікувати помірного тиску хвороб, а якщо воно досягає 10, ми можемо очікувати високого тиску хвороб з боку клімату. Знаючи сприйнятливість сорту та історію поля, ми зможемо прийняти рішення про те, обприскувати чи не обприскувати за слабкого або помірного тиску хвороб у цій ситуації. Накопичене значення 10 може призвести до обприскування на стадії 32 в будь-якому випадку. Рішення про обприскування на більш пізніх стадіях більше залежить від весняного клімату. Якщо ми зможемо накопичити значення ураженості, починаючи зі стадії 10, до значення 6, ми можемо очікувати слабкого тиску хвороби. Якщо це значення досягає 10, ми можемо очікувати помірного тиску хвороби, а якщо це значення досягає 15, ми можемо очікувати високого тиску хвороби, що залежить від кліматичної ситуації.
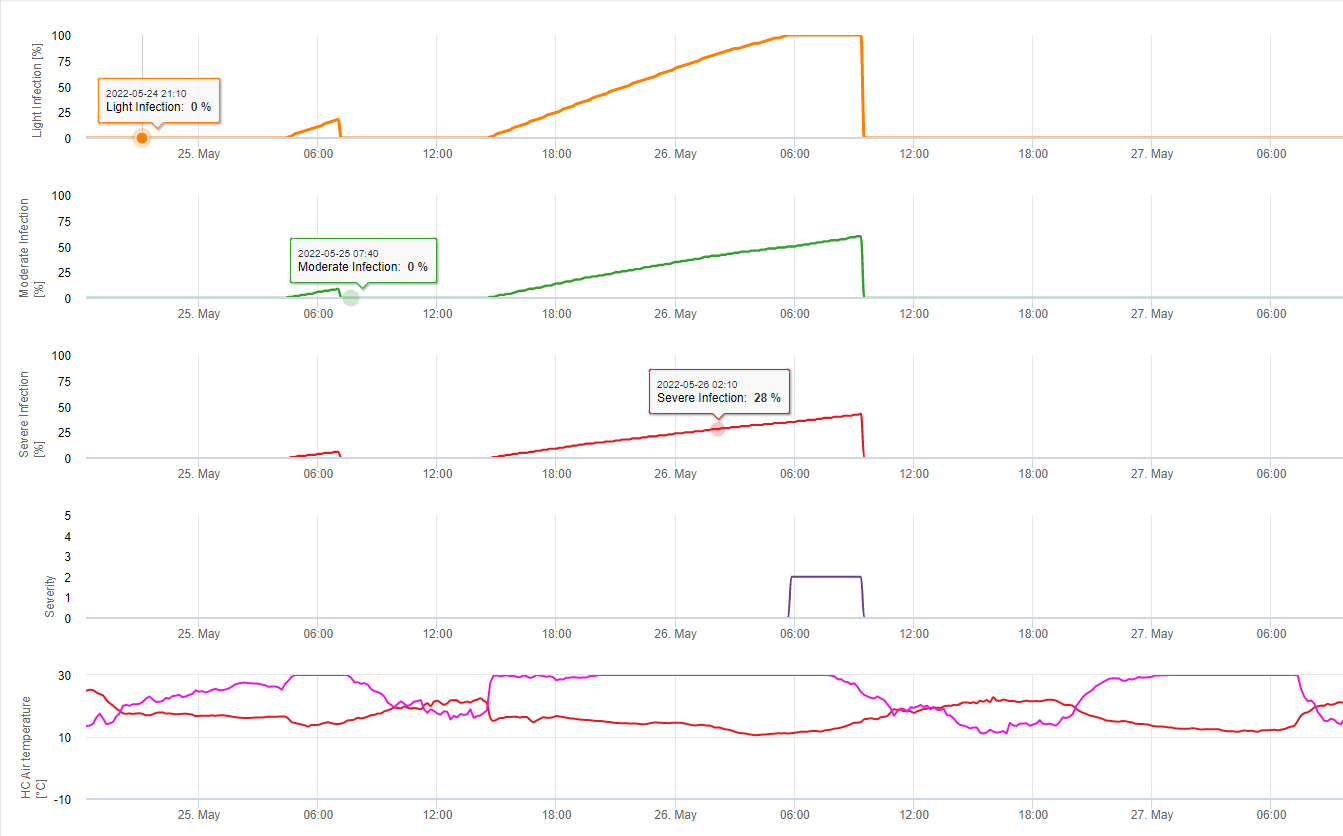
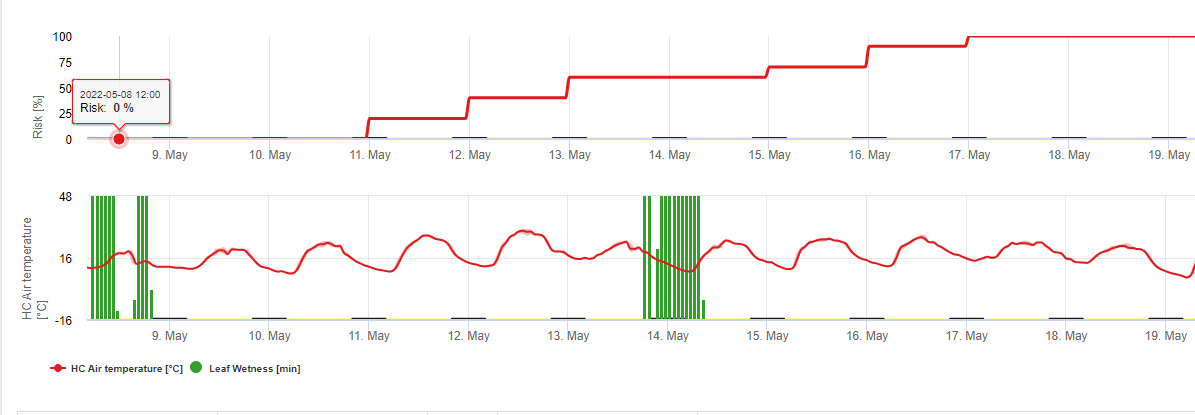
У FieldClimate ми показуємо Septoria tritici Тяжкість разом з трьома різними ступенями тяжкості інфекції на одному графіку (див. вище). Надмірна кількість опадів і тривалий період вологості листя створюють умови для розвитку важкої інфекції через S. tritici були повними 14 та 16 травня. Рівні тяжкості досягли найвищого значення 5 14 травня, що означає високий ризик інфікування.
Біологія інфекції Stagonospora nodorum дещо відрізняється від біології S. tritici, але ця різниця не є достатньо великою для окремої моделі. Тому ми пропонуємо використовувати цю модель для всього комплексу стагноспорозних і септоріозних хвороб зернових культур, включаючи S. passerinii. S. tritici і S. passerinii як правило, потребують більш тривалого періоду зволоження листя, ніж S. nodorum. У районах з високим тиском S. nodorum інфекції, класифіковані як слабкі, що мають ступінь тяжкості 2, слід лікувати більш серйозно, ніж в інших областях.
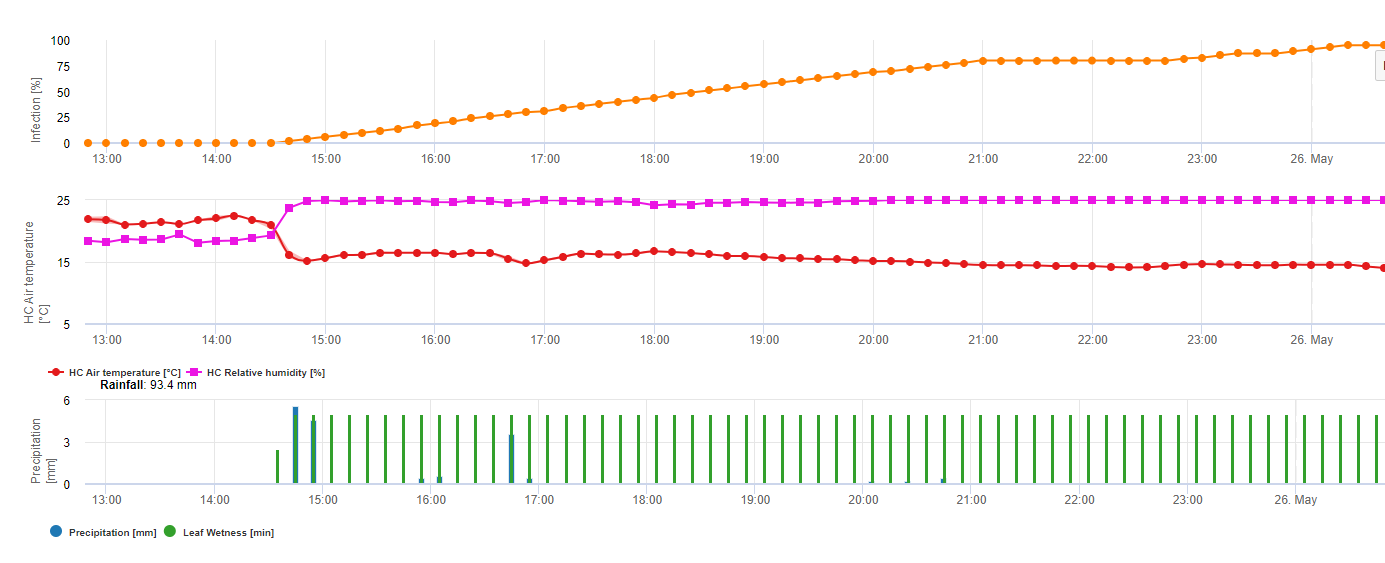
Для Септорія вузлувата (Septoria nodorum) модель ризику показана в FieldClimate (див. вище). Високий ризик був визначений 17 червня та 7 липня (100%). Залежно від сприйнятливої до інфекції стадії рослини, необхідно вжити заходів захисту рослин, якщо ризик досягне 80% (також див. прогноз погоди, захист рослин). Якщо ризик становить 100% і інфекція вже виявлена, для захисту рослин необхідно вжити системних заходів захисту рослин (лікувальне застосування).
Кілька видів рослинних патогенних грибів у роді Коллетотрихум (Colletotrichum) викликають антракноз перцю та багатьох інших овочів і фруктів. До кінця 1990-х років антракноз перцю і томатів був пов'язаний тільки зі стиглими або дозріваючими плодами. З того часу встановилася більш агресивна форма хвороби. Ця форма вражає перець на будь-якій стадії розвитку плодів і може загрожувати прибутковості посівів перцю в районах, де вона закріпилася. Ця хвороба також може вражати помідори, полуницю і, можливо, інші фруктові та овочеві культури.
Симптоми
Кругові або кутові вдавлені ураження розвиваються на незрілих плодах будь-якого розміру. Часто на окремих плодах утворюються множинні ураження. При важкому перебігу хвороби ураження можуть зливатися. Часто на поверхні уражених плодів утворюються концентричні кільця спор гриба від рожевого до помаранчевого кольору. У застарілих ураженнях можна спостерігати чорні структури, які називаються ацервули. Неозброєним оком вони виглядають як маленькі чорні крапки, а під мікроскопом - як пучки крихітних чорних волосків. Патоген швидко і рясно утворює спори і може швидко поширюватися по всій культурі перцю, що призводить до втрати врожаю до 100%. Ураження також може проявлятися на стеблах і листках у вигляді коричневих плям неправильної форми з темно-коричневими краями.
Збудник
Ця форма антракнозу перцю викликається грибком Colletotrichum acutatum. Патоген виживає на рослинних рештках заражених культур та на інших сприйнятливих видах рослин. Гриб не переноситься ґрунтом протягом тривалого часу за відсутності заражених рослинних решток. Гриб також може потрапляти в культуру з зараженим насінням. Під час теплі та вологі періоди, спори розбризкуються дощем або поливною водою від хворого до здорового плоду. Хворі плоди слугують джерелом посівного матеріалу, що дозволяє хворобі поширюватися від рослини до рослини в межах поля. Грибок виживає в насінні та на ньому. Антракноз потрапляє в поле з інфікованими саджанцями або може виживати між сезонами в рослинних рештках чи на бур'янах-хазяїнах. Альтернативними господарями є бур'яни та інші рослини родини пасльонових (помідори, картопля, баклажани), хоча у Флориді інфікування цими рослинами надзвичайно рідкісне. Плоди заражаються, коли спори гриба або заражені рештки потрапляють під час дощу на рослини перцю. Нові спори утворюються в зараженій тканині, а потім поширюються на інші плоди. Працівники також можуть переносити спори з обладнанням або під час роботи з зараженими рослинами. Зараження зазвичай відбувається в теплу, вологу погоду. Температура навколо 80° F (27° C) є оптимальною температурою для розвитку хвороби, хоча інфікування відбувається як при вищих, так і при нижчих температурах. Значні втрати відбуваються під час дощової погоди, оскільки спори змиваються або розбризкуються на інші плоди, що призводить до більшої кількості інфекцій. Хвороба частіше розвивається на зрілих плодах, які тривалий час перебувають на рослині, хоча вона може виникнути як на незрілих, так і на зрілих плодах. Антракноз може розвиватися при температурі від 15°C до 30°C. Але антракноз тривала вологість листя необхідна для задоволення потреб інфекції. При оптимальній температурі від 20°C до 25°C ще 12 годин вологості листя необхідні. Вища або холодніша температура потребує ще довших періодів вологості листя (для розрахунку не потрібна лінійна функція/масив). FieldClimate розраховує можливі події зараження на основі вологості листя та температури під час цієї події.
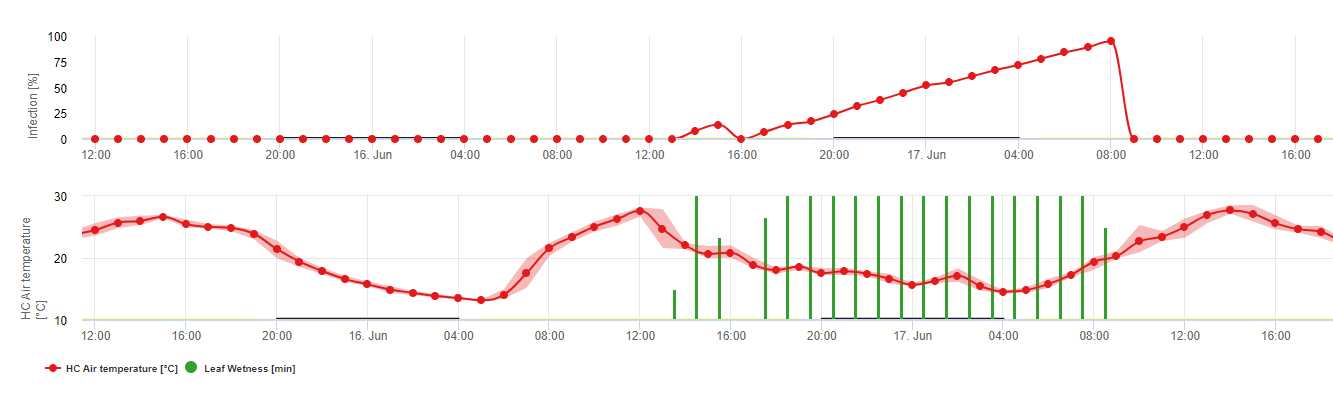
Біологія Plasmopara viticola (несправжня борошниста роса)
P. viticola є облігатним паразитом. Це означає, що для росту потрібні зелені, свіжі органи виноградної лози. У вільний від вегетації період він зберігається, утворюючи плодові тіла, так звані ооспори. Ооспори ооміцетів можуть виживати в ґрунті дуже довго. Тому ми можемо знайти несправжню борошнисту росу в місцях, де зараження можливе не кожного року. Навесні, коли верхній шар ґрунту достатньо вологий і теплий, ооспори утворюють так звані макроспорангії, які можуть випускати до 200 зооспор у вільну воду. Зооспори переносяться вітром у краплинах води на листя та грона. Вони мають два джгутики і рухаються у водяній плівці на нижньому боці листків, грон та молодих ягід, щоб знайти отвір для проникнення в тканину рослини. Вони проникають і проростають у продихи, в які вони переносять всю свою плазму протягом менш ніж однієї години. У мікроскопічних дослідженнях знаходження стоми, інцистація та проростання в стому завершувалися протягом 90 хвилин.
P. viticola росте в міжклітинному просторі і живиться гаусторіями, що проникають в епідермічні та паренхімні клітини. Залежно від температури і відносної вологості він розвиває достатній міжклітинний ріст з достатньою кількістю гаусторій, щоб сформувати субстратне тіло, яке заповнює всю площу субстрату і піднімає епідермальну тканину від паренхіматозної тканини. Це призводить до появи видимого симптому масляної плями.
Ооміцети - це спороношення за відсутності світла при відносній вологості дуже висока. У P. viticola спороношення не відбувається, якщо температура нижче 12°C і відносна вологість нижче 95%. Спорангіафори утворюються з пухирців субстоми і виходять зі стоми. Свіжоутворені спорангії є липкими і можуть бути видалені зі спорангіафорів лише за допомогою води. При зниженні відносної вологості повітря спорангії стають липкими і можуть бути видалені також вітром.
Спорангій випустить у вільний простір до 20 зооспор. Ці зооспори повинні поширюватися вітром у краплях води, щоб потрапити на свіже листя, або ж спорангії можуть поширюватися дощем чи вітром самі по собі. Процес зараження при первинному і вторинному зараженні однаковий.
Враховуючи велике значення статевої стадії для перезимівлі патогена, можна припустити, що типи спарювання придатні для всіх виноградників, де зустрічається несправжня борошниста роса виноградної лози. Формування зооспор відбувається на старому листі протягом літа і ранньої осені.
Первинна інфекція в деталях
Інфекції, що походять від ооспор, називаються первинними інфекціями. Цей термін вводить в оману, оскільки на початку літа може виникнути кілька первинних інфекцій. В епідеміологічних подіях первинна інфекція не відіграє важливої ролі, якщо на виноградниках є достатня кількість олійних плям і інфекційний потенціал літніх спор (спорангіїв) перевищує ооспори.
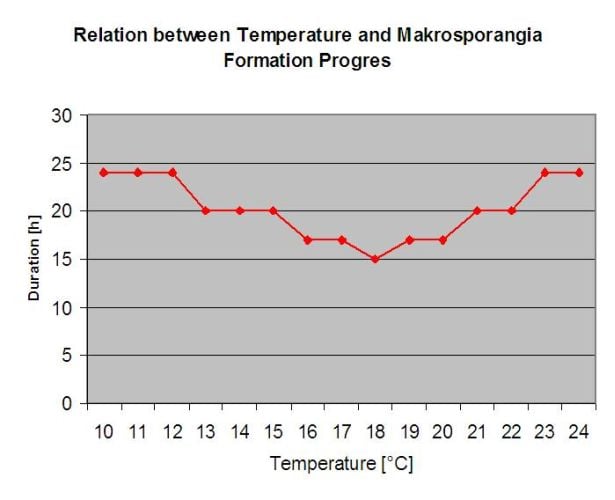
Зимуючі ооспори розвиваються у так звані макроспорангії за достатньої відносної вологості повітря протягом 24 годин. Макроспорангії вивільняють зооспори у вільну воду та під час сильного дощу (наприклад, під час грози). Спори переносяться на листя виноградної лози та/або зелені пагони. Таким чином відбувається первинне зараження, Потрібні довші дощі дощові періоди або кілька гроз, що йдуть підряд. Перший дощ призводить до того, що опале листя насичується водою, а сильні дощі в наступні дні змушують макроспорангії вивільняти зооспори, які потрапляють на листя або пагони винограду. Для успішного зараження необхідний досить тривалий період зволоження листя, щоб зооспори змогли досягти продихів листя або пагонів і заразити їх (на ілюстрації вище показано цикл розвитку несправжньої борошнистої роси виноградної лози (Plasmopara viticola).
Модель первинної інфекції несправжньої борошнистої роси спочатку перевіряє, чи сприятлива погода для розвитку макроспорангіїв. Це відбувається до тих пір, поки листя вологе, або відносна вологість після дощу не опускається нижче 70%. Залежно від температури, макроспорангії можуть розвиватися протягом 16-24 годин. Якщо наявні зрілі макроспорангії, це буде показано графічно на дисплеї первинної інфекції несправжньої борошнистої роси. Якщо макроспорангії присутні, сильний дощ може сприяти поширенню їх зооспор. Безперервний дощ 5 мм інтерпретується як сильний дощ, і зооспори поширюються, первинне зараження можливе, якщо вологість листя триває досить довго.
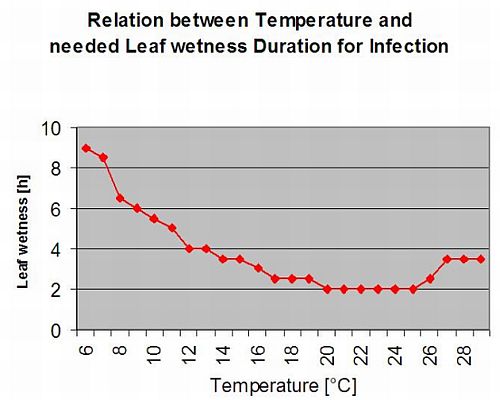
Вторинні інфекції
Вторинні інфекції Plasmopara viticola можливе лише тоді, коли на вашому винограднику або поблизу нього вже є зрілі олійні плями. Олійні плями є зрілими, коли вони здатні до спороношення (утворення спорангіїв). Спорангії виробляються тільки ніч. Сонячне світло пригнічує спороношення. Спорангії утворюються, якщо це тепліше 12°C і відносна вологість дуже висока. Швидкість утворення спорангіїв зростає з підвищенням температури до 24°C. Оптимальна температура для спороношення на європейських сортах винограду (Vitis vinifera) становить близько 24°C. Якщо температура перевищує 29°C, спороношення не відбувається. У нашій моделі ми перевіряємо, чи вологість понад 95% відбувається вночі. Якщо такий стан триває протягом години, коли накопичена температура перевищує 50°C, спороношення завершується, і нові спорангії Плазмопара існують на винограднику. Наприклад, години при 50°C відповідають 4 годинам при 13°C або 3 годинам при 17°C.
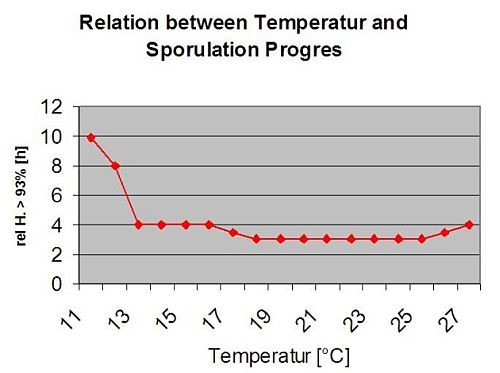
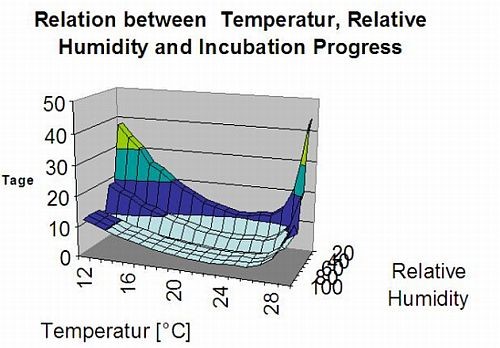
На наступному графіку показано зв'язок між температурою та умовами вологості (відносною вологістю, вологістю листя, опадами), які використовуються для моделювання інфекційних подій в fieldclimate.com.
Борошниста роса томатів може бути викликана трьома збудниками у всьому світі.
Leveillula taurica (Oidiopsis taurica) є патогеном широкого спектру видів хазяїв у теплому посушливому та напівпосушливому кліматі в Азії, Середземномор'ї, Африці, а віднедавна і на південному заході Сполучених Штатів Америки.
Erysiphe orontii (E. cichoracearum і E. polyphaga) - ще один вид, поширений серед багатьох рослин-господарів як у помірних, так і в тропічних регіонах.
І як третій вид Odium lycopersicum.
Фактори розвитку захворювання
- рівень відносної вологості > 50% (оптимальний RH > 90%)
- вільна вода на поверхні листя не потрібна
- температурний діапазон: 10-35 °C (найкраще нижче 30 °C)
Борошниста роса - це хвороба, що спричиняється інокулятом. Тому можна визначити лише ризиковані періоди, основним фактором ураження, наприклад, спалаху, є початковий інокулят (який активний у широкому діапазоні температур). Отже, для стратегій контролю: поєднуйте моделювання ризикованого періоду з моніторингом грибкового інокуляту (хвороби) в полі!
Томат відкритого ґрунту
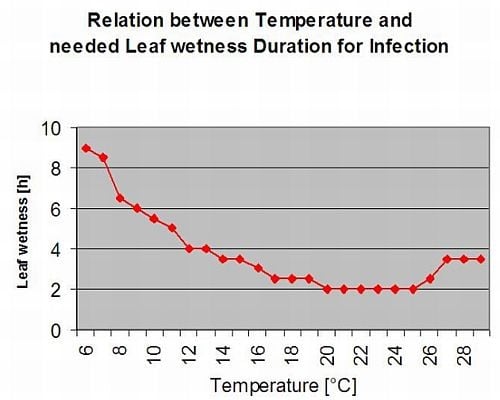
Іржа спаржі Спаржа пуччінія сприяє вологий і теплий клімат. Невелика кількість дощу і вітер розподіляють уредієспори, тоді як сильні дощі можуть змити їх у ґрунт. З цієї причини період вологості листя, коли випадає більше 10 мм опадів, не вважається інфекційним періодом. Періоди вологості листя під час невеликих дощів та роси тривалістю від 3 до 8 годин при температурі від 25°C до 10°C можуть бути періодами інфікування уредіаспор влітку та наприкінці літа.
Ця модель оцінює дату першої появи C. beticola на основі накопичених середньодобових температур з першого січня понад 5°C. Для періодів з відносною вологістю вище 60% приймається поріг 1006,2°C, а за відсутності відносної вологості діє поріг 1081,9°C.
Ця модель була розроблена для Італії і використовується в Італії та Німеччині.
Для 2010 року в Штирії CercoPrim вказує на 19 червня як дату першого обприскування. Це приблизно та сама дата, яку модель Pessl Instruments Risk або модель DIV вказали б для обприскування сприйнятливих до вовчка сортів.
Збудник
Пурпурова плямистість на спаржевих списах і папороті викликається грибком Stemphylium vesicarium. Гриб переживає зиму як статеві спори (аскоспори) в мішечку (аскусі), що утворюються в зимуючих структурах (псевдотеціях), які виглядають як маленькі чорні крапки на залишках спаржі з попереднього сезону. У цьому випадку, як і в попередньому випадку. аскоспори вивільняються з асцису під час дощу і можуть переноситися вітром на нові рослини спаржі, де вони спричиняють первинне інфікування в період вегетації. Ці нові інфекції призводять до утворення спор (конідій), що утворюються безстатевим шляхом, які, в свою чергу, можуть викликати вторинні інфекції, і цей процес повторюється до тих пір, поки температура і кількість опадів є сприятливими.
Хвороба
Поява пурпурової плямистості як значної проблеми у виробництві спаржі в Мічигані може бути пов'язана з прийняттям системи нульового обробітку ґрунту, при якій суху папороть попереднього сезону подрібнюють у квітні і залишають на поверхні ґрунту. Ці залишки зберігаються протягом сезону збору врожаю (середина червня) і є видимими протягом періоду росту папороті (кінець червня - вересень) і є джерелом аскоспор, які починають інфікування на початку вегетаційного періоду. Симптоми хвороби такі: Хвороба проявляється у вигляді численних, злегка заглиблених, пурпурових плям з коричневими центрами на зібраних списах і папороті. Ураження на списах часто виявляють з навітряного боку, оскільки пісок, що розноситься вітром, спричиняє рани, які сприяють інфікуванню. В епідемічні роки плямистість може з'являтися на 60-90% списів і може призвести до відбраковування врожаю, особливо для продажу на свіжому ринку. Плямистість також з'являється на спаржевій папороті, вражаючи головне стебло, вторинні гілки та голки (кладофіли). Сильне зараження папороті може призвести до передчасної дефоліації рослини. Зростання тяжкості хвороби пурпурової плямистості пов'язане з тривалими періодами дощів, туману або роси.
Автор тексту: Мері К. Хаусбек, професор і спеціаліст з питань поширення знань, Університет штату Мічиган, кафедра патології рослин, Е. Лансінг
TomCast для спаржі
Передісторія: TOMCAST (TOMato disease foreCASTing) - це комп'ютерна модель на основі польових даних, яка намагається передбачити розвиток грибкових захворювань, а саме Ранній фітофтороз, септоріоз листя та антракноз на томатах. Польові реєстратори даних щогодини записують дані про вологість і температуру листя. Ці дані аналізуються протягом 24 годин і можуть призвести до формування показника тяжкості хвороби (Disease Severity Value, DSV), який, по суті, відображає приріст розвитку хвороби. По мірі накопичення DSV, тиск хвороби на культуру продовжує зростати. Коли кількість накопичених DSV перевищує інтервал між обприскуваннями, рекомендується застосування фунгіциду для зменшення тиску хвороби.
TOMCAST походить від оригінальної моделі F.A.S.T. (Forecasting Alternaria solani on Tomatoes), розробленої докторами Медденом, Пенніпекером і Макнабом в Університеті штату Пенсильванія (PSU). Модель PSU F.A.S.T. була в подальшому модифікована доктором Пітбладо в коледжі Ріджтаун в Онтаріо в те, що ми зараз визнаємо як модель TOMCAST, яка використовується відділом розвитку Університету штату Огайо.
DSV є: Значення тяжкості хвороби (DSV) - це одиниця виміру, що надається конкретному приросту розвитку хвороби (раннього фітофторозу).
Іншими словами, DSV - це числове представлення того, наскільки швидко або повільно хвороба (ранній фітофтороз) накопичується на томатному полі. DSV визначається двома факторами: вологістю листя і температурою в години "мокрого листя". Зі збільшенням кількості годин з мокрим листям і температури DSV накопичується швидше. Нижче наведено діаграму "Тяжкість хвороби".
І навпаки, за меншої кількості годин зволоження листя та нижчої температури DSV накопичуються повільніше, якщо взагалі накопичуються. Коли загальна кількість накопичених DSV перевищує встановлену межу, яка називається інтервалом або порогом обприскування, рекомендується обприскування фунгіцидом, щоб захистити листя і плоди від розвитку хвороб.
Інтервал між обприскуваннями (який визначає, коли слід обприскувати) може коливатися в межах 15-20 DSV. Точне значення DSV, яке повинен використовувати виробник, зазвичай надається переробником і залежить від якості плодів і кінцевого використання томатів. Дотримання інтервалу обприскування 15 DSV є консервативним використанням системи TOMCAST, тобто ви будете обприскувати частіше, ніж фермер, який використовує інтервал обприскування 19 DSV з системою TOMCAST. Компроміс полягає в кількості обприскувань протягом сезону і потенційній різниці в якості плодів.
 TomCast зі спаржею
TomCast зі спаржею
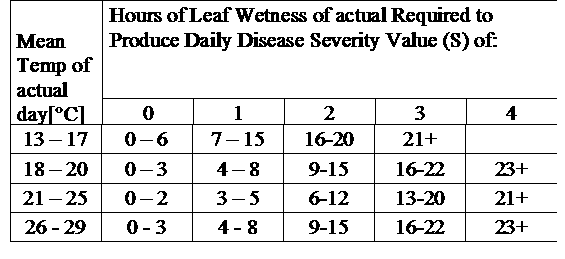
Дослідження визначили, що прогнозатор хвороб Tom-Cast є перспективною альтернативою календарному обприскуванню папороті на комерційних полях спаржі. Tom-Cast попереджає виробників, коли умови навколишнього середовища сприятливі для розвитку хвороби пурпурової плямистості (тривалі росяні або дощові періоди, що супроводжуються теплою температурою). Ефективні фунгіциди, що застосовуються відповідно до прогнозу хвороб Tom-Cast, дозволяють виробникам боротися з пурпуровою плямистістю спаржі, заощаджуючи при цьому гроші та зберігаючи навколишнє середовище.
Текстовий залік: Джим Ясінські, координатор TOMCAST в Огайо, Індіані та Мічигані
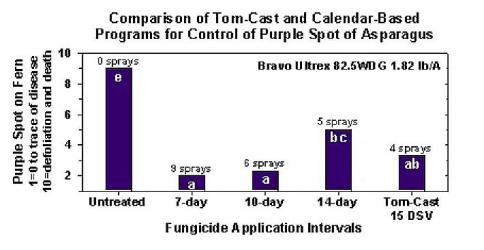
Модель інфекції пурпурової плямистості
У "The Stemphylium versicarium Модель інфекції базується на роботах SUHERI та PRICE на цибулі та LLorent, VILARDELL, BUGIANI, GHERARDI та MONTESINO на груші.
Розраховано криві інфікування для легких, середніх та важких інфекцій.
Завдяки такому поділу на сприятливі, помірно сприятливі та дуже сприятливі ситуації, фермер сам вирішує, наскільки сильним буде тиск хвороб на конкретному полі і чи потрібно йому боротися з конкретною інфекцією.
Умови:
Температура: 11-30°C
: > 0 (старт для інфекції) або відносної вологості > 90% (тільки якщо інфекція вже була до розрахунку і все ще тримається)
легка інфекція (11-30°C), помірна інфекція (13-30°C), сильна інфекція (16-30°C), масив з тривалістю вологості листя (див. вище)
Скидання: якщо відносна вологість нижче 70%.