Soya Pası, özellikle Asya soya pası, mantar patojeninin neden olduğu ciddi bir soya fasulyesi hastalığıdır Phakopsora pachyrhizi. Hastalığın dağılımı: Asya ve Avustralya ve son zamanlarda Afrika, Kuzey Amerika ve Güney Amerika. Hastalık ilk olarak 1902 yılında Japonya'da görülmüştür. Patojen Amerika üzerinden Asya ve Avustralya'ya yayılmıştır. Soya pası dondurucu soğukların olduğu bölgelerde kışı geçiremez, ancak rüzgarla o kadar büyük mesafelere hızla yayılabilir, gelişimi o kadar patlayıcı olabilir ve o kadar hızlı yaprak kaybına neden olabilir ki, şu anda dünyanın soya fasulyesi yetiştirilen bölgelerinde en korkulan hastalıklardan biridir.
Belirtiler ve İşaretler
Soya fasulyesi pasının neden olduğu ilk belirtiler Phakopsora pachyrhizi yapraklarda çok küçük kahverengi veya kiremit kırmızısı lekeler olarak başlar. Tarlada, bu lekeler genellikle çiçeklenme sırasında veya sonrasında alt kanopide başlar, ancak fideler belirli koşullar altında enfekte olabilir. Genellikle ilk lezyonlar yaprak sapı ve yaprak damarlarının yakınında yaprakçığın tabanına doğru ortaya çıkar. Yaprakçığın bu kısmı muhtemelen çiği daha uzun süre tutarak koşulları enfeksiyon için daha elverişli hale getirir. Lezyonlar küçük kalır (2-5 mm çapında), ancak hastalık ilerledikçe sayıları artar. Bu lezyonlarda, çoğunlukla alt yaprak yüzeyinde uredinia adı verilen püstüller oluşur ve bunlar çok sayıda urediniospor üretebilir.
Kabarık püstüller, özellikle sporlanma sırasında çıplak gözle görülebilir. Lezyonlar küçük olsa da, her lezyonda genellikle birkaç püstül (uredinia) bulunur. Püstüller aktif olduğunda lezyonlar tamamen urediniosporlarla kaplanabilir. Soya pası urediniosporları soluk sarı-kahverengi ila renksizdir ve ekinülat (kısa dikenler) yüzey süslemesine sahiptir. Bu renklenme, sporları genellikle kırmızımsı kahverengi (pas renkli) olan diğer birçok pas patojeninden farklıdır. Çimlenme P. pachyrhizi urediniosporlar ekvatoral (merkezi) bir gözenek aracılığıyla meydana gelir ve mantarın konağa doğrudan veya bir stoma aracılığıyla nüfuz etmek için kullandığı bir appressorium ile biten bir germ tüpü üretir.
Bir yaprakçık üzerinde giderek daha fazla lezyon oluştukça, etkilenen alan sararmaya başlar ve sonunda yaprakçık bitkiden düşer. Soya fasulyesi pası genellikle alt gölgelikte başlarken, tüm yapraklarda bir miktar hastalık oluşana kadar bitkide hızla ilerler. Ciddi derecede hastalıklı bitkiler tamamen yapraksız kalabilir. Etkili yaprak dokusunun kaybı, hem daha az hem de daha küçük tohumdan kaynaklanan verim düşüşlerine neden olur. Verim kayıplarının 30 ila 80% kadar yüksek olduğu bildirilmiştir, ancak kayıp miktarı hastalığın ne zaman başladığına ve ne kadar hızlı ilerlediğine bağlıdır. Soya fasulyesi pası yaprakların yanı sıra yaprak saplarında, gövdelerde ve hatta kotiledonlarda da görülebilir, ancak pas lezyonlarının çoğu yapraklarda görülür.
Lezyonlar ten rengi ya da kırmızı-kahverengi olabilir. Bronz lezyonlarda çok sayıda urediniospor üreten çok sayıda püstül bulunur. Orta derecede direnç reaksiyonu olduğu düşünülen kırmızı-kahverengi lezyonlar, sadece birkaç urediniospor üreten birkaç püstüle sahiptir. Hastalık Yönetimi bölümünde tartışılacağı gibi, bu lezyon tipi patojenin türüne bağlıdır ve taba rengi lezyonlarla aynı yaprakta görülebilir veya taba rengi lezyonlar yaşla birlikte kırmızı-kahverengiye dönüşebilir. Kudzu gibi diğer konukçulardaki belirti ve bulgular benzerdir, ancak lezyon boyutu farklılık gösterebilir. Püstüller yaşlandıkça siyaha dönüşebilir. Bunun nedeni püstüllerde bir teliospor tabakasının oluşması ve püstüllerin ürinia'dan telia'ya dönüşmesidir.
Teliosporların iki işlevi vardır: canlı bir konakçının yokluğunda mantarın hayatta kalması (overseasoning) ve eşeyli üreme. Teliosporların kalın duvarları mantarı çevreden ve diğer organizmaların saldırısından korur. Paslarda, teliosporlar çimlenerek bir basidium ve dört basidiospor oluşturur ve bu sırada eşeyli rekombinasyon meydana gelir. Çimlenme P. pachyrhizi teliosporlar sadece laboratuvarda gözlemlenmiştir ve bu hastalığın sahada devam etmesine önemli bir katkıda bulunmuyor gibi görünmektedir.
Patojen Biyolojisi
Soya fasulyesinde pasa neden olan iki yakın akraba mantar vardır: Phakopsora pachyrhizibazen Asya veya Avustralya soya fasulyesi pas patojeni olarak adlandırılan, ancak artık batı yarımkürede de görülen ve P. meibomiaesadece batı yarımkürede bulunan Yeni Dünya soya fasulyesi pas patojeni olarak adlandırılır. Birkaç küçük özellik dışında, iki mantar morfolojik olarak aynı görünür, ancak P. pachyrhizi soya fasulyesi üzerinde çok daha agresiftir. P. meibomiae. Bugüne kadar, P. meibomiae Orta ve Güney Amerika'da önemli verim kayıplarına neden olduğu belgelenmemiştir. İki tür DNA analiz protokolleri kullanılarak ayırt edilebilir.
Diğer paslar gibi soya fasulyesi pası patojenleri de büyümek ve çoğalmak için canlı bir konukçuya ihtiyaç duyan zorunlu parazitlerdir. Doğal koşullar altında konukçudan uzakta urediniospor olarak sadece birkaç gün hayatta kalabilirler. Her iki soya fasulyesi pas patojeni de, bildiğimiz kadarıyla, sadece iki tür spor üretir: urediniosporlar ve teliosporlar (Şekil 15). Bu durum, beş spor aşamasına kadar sahip olabilen diğer paslarla (örneğin buğday sap pası) tezat oluşturmaktadır. Soya fasulyesi pası için, çoğu pas gibi, üredinial aşama tekrar eden aşamadır. Bu, ürediniosporların üretildikleri aynı konukçuyu (soya fasulyesi) aynı mevsim boyunca enfekte edebileceği anlamına gelir. Salgınlar sadece birkaç püstülden hızlı bir şekilde gelişebilir çünkü spor üreten püstüller enfeksiyondan sonra 7 ila 10 gün gibi kısa bir sürede üretilir ve her püstül yüzlerce urediniospor üretebilir. Teliosporlar eski lezyonlarda üretilir, ancak doğada çimleniyor gibi görünmezler ve alternatif konak, aecia veya spermogonia bilinmemektedir.
Teliosporların çimlenmesi olmadan eşeyli üreme gerçekleşemez. Eşeyli üremenin olmaması, pas mantarının değişkenliğini sınırlamalıdır, ancak yine de pas mantarının türlerinde önemli bir değişkenlik vardır. P. pachyrhizi virülans ile ilgili olarak. Bu durum soya fasulyesinde direnç için tek genlerin kullanımını sınırlamıştır, çünkü kısa sürede direnç geninin üstesinden gelen yeni patojen izolatları ortaya çıkmaktadır. Bu değişkenliğin nasıl ortaya çıktığı bilinmemektedir. P. pachyrhizi. Buğday şerit pası, Puccinia striiformisile benzer bir yaşam döngüsüne sahiptir. P. pachyrhizi İşleyen bir telial aşaması ve dolayısıyla eşeyli üremesi yoktur, ancak birçok ırkı vardır. Her bir direnç geni o kadar spesifik olabilir ki, mantarın doğru genindeki tek bir mutasyon, yeni direnç genine sahip konakçılarda virülan olmasını sağlar.
Epidemiyoloji
Soya fasulyesi pas salgınları havadaki inokulumların (urediniosporlar) gelişiyle başlar. Bu patojen paslar arasında benzersizdir çünkü inokulum kaynağı olarak hizmet edebilecek birçok alternatif konukçuya sahiptir. Alternatif konukçular, aynı patojenle enfekte olabilen ancak patojenin yaşam döngüsünü tamamlamak için gerekli olmayan diğer bitkilerdir. Alternatif konukçular, bir patojenin yaşam döngüsünü tamamlaması için gerekli olan ana konukçudan başka bir bitki olan alternatif konukçu ile karıştırılmamalıdır. Güney Amerika, Orta Amerika, Karayip havzası, güney Teksas ve Florida gibi don olmayan bölgelerde, inokulum kaynağı yakınlardaki gönüllü soya fasulyesi bitkileri, kudzu veya başka bir alternatif konukçu olabilir. Orta Batı Amerika Birleşik Devletleri gibi don yaşanan bölgelerde, inokulum yüzlerce mil uzakta olabilecek kışlama kaynaklarından üflenmelidir.
Zorunlu patojenlerin uzak bir bölgeye yeniden sokulması, buğday kök pası ve tüylü mildiyöler, örneğin tütündeki mavi küf gibi diğer birçok hastalıkta meydana gelir. Çünkü sporları P. pachyrhizi ultraviyole radyasyona duyarlı olduğundan, bu pas sporlarının uzun mesafeli hareketi muhtemelen bulutların sporları güneşten koruduğu fırtına sistemlerinde gerçekleşir. Canlı sporlar uygun bir konağın yaprak yüzeyine indiğinde, enfeksiyon ve ardından salgın gelişimi çevresel koşullara bağlıdır. Genel olarak enfeksiyon, yapraklar ıslak olduğunda ve sıcaklık 8°C ile 28°C arasında olduğunda meydana gelir; optimum sıcaklık 16°C ile 28°C arasındadır. 25°C'de, yaprak ıslaklığının 6 saat gibi kısa bir süresinde bazı enfeksiyonlar meydana gelir, ancak 12 saat en uygunudur. Enfeksiyondan sonra, urediniosporlu lezyonlar ve püstüller 7 veya 8 gün içinde ortaya çıkabilir ve bir sonraki enfeksiyon döngüsü başlar.
Bu kısa yaşam döngüsü, doğru koşullar altında soya pası salgınlarının neredeyse tespit edilemeyen seviyelerden çok yüksek seviyelere hızlı bir şekilde çıkabileceği anlamına gelir. Soya pası salgınları, tespit edilebilir seviyelerin altından bir ay içinde yaprak dökmeye kadar ilerleyebilir. Salgınlar bundan daha da hızlı ilerliyor gibi görünebilir, çünkü erken enfeksiyonlar alt gölgelikte meydana gelir ve bulunması zordur. Çevrenin yanı sıra, bitki yaşı da soya fasulyesi pas salgınlarını etkiler. Genellikle, sezonun başlarında yüksek inokulum seviyeleri olmadıkça, çiçeklenmeye kadar soya fasulyesinde pas lezyonları bulunmaz. Bunun nedeni, konukçu üreme aşamalarına girdikçe bitkilerin pasa karşı daha duyarlı olması, kanopinin alt kısımlarında sporların UV radyasyonundan daha fazla korunması veya kanopi kapandıkça kanopideki koşulların daha nemli hale gelmesi olabilir. Her durumda, lezyonlar herhangi bir büyüme aşamasında oluşabilir, ancak hastalıktaki büyük artışlar çiçeklenme sonrasına kadar gerçekleşmez.
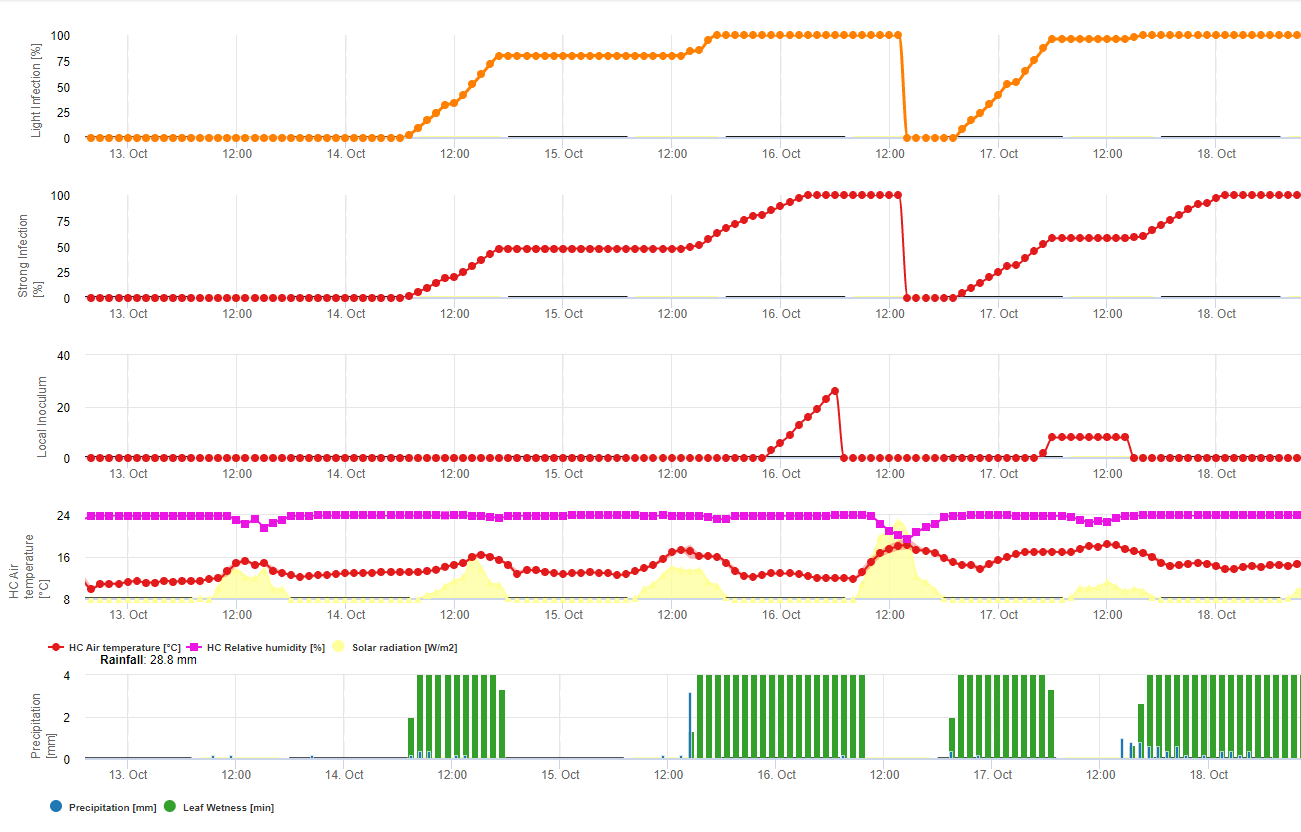
Soya fasulyesi pası için iki model FieldClimate'de tanımlanmış ve hesaplanmıştır:
Model 1: Soya fasulyesi pasının enfeksiyon ve şiddetinin hesaplanması için ana faktörler yaprak ıslaklığı ve sıcaklıktır (şiddet oranları (1-3 arası) hesaplanmaya başlayana kadar enfeksiyonun 100% olmasına dikkat edin) Yapraklar ıslak olmalı ve sıcaklık 8 ila 28°C arasında olmalıdır - enfeksiyon hesaplanmaya başlar (7200 °dakika toplamı 100%'dir). Enfeksiyon 100% ise ve sıcaklık ve yaprak ıslaklık süresine bağlı olarak 1-3 arası şiddet sınıfları hesaplanmaya başlar.
Model 2: Burada Soya fasulyesi pası hesaplamasına çok daha fazla faktör dahil edilmiştir: Sıcaklık, Yaprak ıslaklığı, bağıl Nem, Yağış ve güneş radyasyonu (mantar hifleri gündüz vakti stomalara girebilir).
Soya pası enfeksiyonu 100%'ye ulaştığında şiddet hesaplanmaya başlar. Her ikisi de (enfeksiyon ve şiddet 100% ise) yerel inokulumun oluşumu (kurulması?) tahmin edilir.
Edebiyat:
- Ivancovich, A. y Botta, G. 2002. La roya de la sojo en la Argentinia. EEA Pergamino. RevistadeTEchnologia, Agropecuaria Vol. 7 (21), pp 16-17.
- Melching, J. S., Dowler, W. M., Koogle, D. L. ve Royer, M. H. 1989. Yaprak ıslaklık dönemlerinin süresi, sıklığı ve sıcaklığının soya fasulyesi pası üzerindeki etkileri. Plant Dis. 73:117-122.
- X.B. Yang, E.M. Del Ponte ve A.P. Dias. Brezilya ile karşılaştırarak soya fasulyesi pas riskini bilmek. Bitki Patolojisi Bölümü, Iowa Eyalet Üniversitesi
- Yang, X. B. 1995. Soya fasulyesi pası riskinin değerlendirilmesi ve yönetimi. Soya pası çalıştayı bildirileri, 9-11 Ağustos 1995. J. B. Sinclair ve G. L. Hartman, eds. Ulusal Soya Araştırma Laboratuvarı, Urbana, IL
Sclerotinia kök çürüklüğü veya soya fasulyesinin beyaz küfü, mantarın neden olduğu bir hastalıktır Sclerotinia sclerotiorum. Hastalık, soya fasulyesi istila edilmiş toprağa ekildiğinde ve uzun süreli yağışlı havalarda yoğun bir bitki örtüsü olduğunda tohum veriminde büyük düşüşlere neden olabilir veya hatta bir ürünü tamamen yok edebilir. Verim kayıpları genellikle hastalık insidansı 15 % veya daha fazla olduğunda meydana gelir. Kuzey Dakota'daki çoğu yılda, Sclerotinia kök çürüklüğü yetiştiriciler için yalnızca küçük ila orta dereceli bir sorundur ve hastalık kuraklık yıllarında nadiren görülür. Yağışlı hava, hastalık gelişiminde önemli bir faktördür. Sulanan soya fasulyelerinde kalıcı bir sorun olmuştur. Tohum verimindeki azalmanın yanı sıra, hastalık tohum kalitesinin düşmesine ve tohumun mantarın siyah sklerotisi ile kirlenmesine de neden olur. Tohum kontaminasyonu ihraç edilen tohumlar için ciddi bir sorun olabilir çünkü yabancı giriş limanlarında tohum partisinin reddedilmesine neden olabilir. Ayrıca, toprağa geri dönen sklerotia, rotasyondaki diğer ürünleri de etkileyebilir. Sclerotinia kök çürüklüğü, yetiştiriciler tarafından patojen ve hastalık döngüsünün anlaşılmasıyla yönetilebilir.
Semptomlar
Belirtiler genellikle sıralar arasındaki ürün örtüsü kapanana ve nemli bir mikro iklim yaratana kadar gözlenmez. Yaprakların solması ve soldurulmasının ardından bitkilerin ölmesi genellikle gözlenen ilk belirtilerdir. Bitki örtüsü altında yapılacak yakından bir inceleme, saplar, yapraklar veya baklalar üzerinde pamuksu, beyaz miselyal (mantar iplikleri) bir büyüme ortaya çıkaracaktır. Lezyonlar ana gövdelerde ve yan dallarda gelişir. Sonunda lezyonlar sapları sarar ve üstteki bitki parçaları ölür. Gövdeler ağarmış ve bazen ileri çürüme nedeniyle parçalanmış görünür. Bitki dokusu üzerinde büyüyen beyaz miselyumdan çeşitli şekil ve boyutlarda büyük, siyah sklerotia oluşacaktır. Sklerotia ayrıca gövde özünde de oluşur ve karakteristik silindirik bir şekle sahiptir. Hastalıklı baklalardaki tohumlar genellikle buruşur ve mantar tarafından enfekte olabilir veya siyah sklerotia ile yer değiştirebilir. Enfekte bitkiler hasat edildiğinde tohum genellikle sklerotia ile kontamine olur.
Patojen Biyolojisi
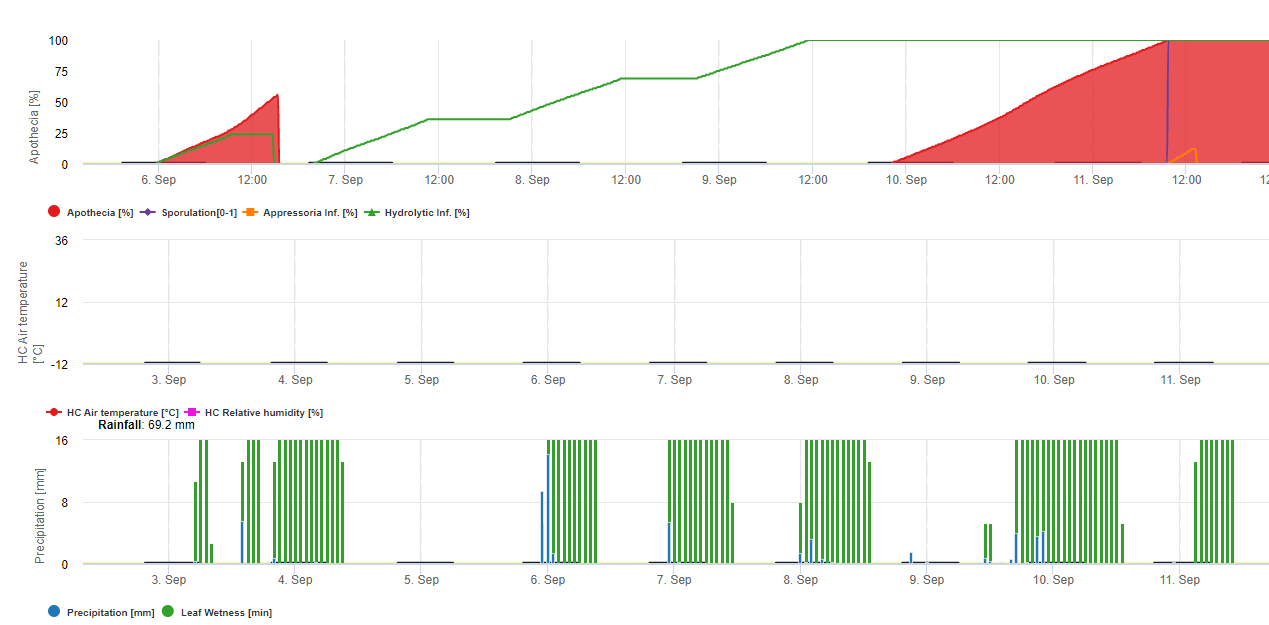
Fungus 370'in üzerinde bitki türünden oluşan geniş bir konukçu yelpazesine sahiptir ve ayçiçeği, kuru fasulye, kanola (kolza), patates, yonca, karabuğday, acı bakla, hardal, yer elması, aspir, mercimek, keten, bezelye ve birçok sebze gibi çok çeşitli ürünlerde hastalıklara neden olur. Kuzey Dakota'da bu patojenin keten ve patates gibi bazı ürünlerde ciddi hasara neden olması nadirdir. Ayrıca bataklık mürver, kuzu otu, domuz otu, Kanada devedikeni ve yabani hardal gibi birçok yaygın geniş yapraklı yabani ot konukçuları da vardır. Soya fasulyesinde beyaz küfe neden olan mantar, ayçiçeği, kuru fasulye, kanola ve diğer ürünlerde beyaz küf veya Sclerotinia hastalığına neden olan mantarla aynıdır. Sclerotinia sclerotiorum esas olarak toprakta sklerotia olarak kışı geçirir. Sklerotia filizlenerek apothecia adı verilen (yaklaşık sekizde bir ila dörtte bir inç çapında) küçük ten rengi ila kahverengi mantarlar oluşturur. Bunlar, soya fasulyesi ve diğer duyarlı ürünlerde hastalığı başlatan askospor olarak adlandırılan sporlar üretir.
Hastalık Döngüsü
Nem ve çiçeklenme hastalık gelişiminde kritik faktörlerdir. Hastalık genellikle ürün örtüsü kapanmadan önce ortaya çıkmaz, çünkü yoğun bir örtü serin sıcaklıkları ve gövdelerin etrafında nemli bir mikro iklimi destekler ve yağmur veya sulamayı takiben yüksek toprak nemini korur. Hastalığın başlaması da çiçeklenme ile yakından ilişkilidir. Yedi ila 14 günlük yüksek toprak nemini takiben, toprağın üst birkaç santimi içindeki sklerotia, mantar benzeri apothecia'yı oluşturmak için çimlenecektir. Tek bir sklerotium birkaç apothecia üretebilir. Apothecia, askosporlarını zorla havaya fırlatacak ve burada hava akımları ile soya fasulyesi bitkilerine taşınacaktır. Askosporların en önemli kaynağı tarla içinde üretilen apothecia'lardır, ancak askosporlar komşu veya yakın tarlalardan da gelebilir. Bir apothecium birkaç gün içinde çok büyük miktarlarda askospor üretebilir. Askosporlar bitki dokusu üzerinde kısa sürelerle hayatta kalır ancak kışı geçirmez. Askosporlar bitkiyi enfekte etmeden önce çimlenmek ve büyümek için bir su tabakasına ve ölü veya yaşlanan çiçek dokuları gibi bir besin tabanına ihtiyaç duyar. Çiçek dokusu enfeksiyonları başlatmak için en önemli besin kaynağıdır. Enfeksiyonlar genellikle yaşlanan çiçek dokularının düşüp yerleştiği gövde akslarında başlar. Enfeksiyonlar dolu veya diğer yaralanmaların neden olduğu yaralar yoluyla da meydana gelebilir. Bitki yüzeyindeki su tabakası lezyonların gelişimini teşvik eder ve doku hasarı miktarını artırır. İlk hastalık gelişimi genellikle 40 saatten fazla sürekli bitki yüzeyi ıslaklığı gerektirir, ancak hastalık bir kez başladığında, daha kısa ıslaklık süreleri lezyon gelişimine izin verecektir. Hastalığın uzun süreli bulutlu, nemli, yağmurlu havalarla ilişkilendirilmesinin nedeni budur. Bitki yüzeyleri kurudukça hastalığın ilerlemesi yavaşlar. 59 ila 75 Fahrenheit derece arasındaki serin sıcaklıklar hastalık gelişimini destekler. Bitki örtüsünün yoğunluğu arttıkça çevresel koşullar hastalık için daha elverişli hale gelir. Katı tohumlanmış soya fasulyesi ve yatık ekinler bu nedenle hastalık gelişimine daha yatkındır. Miselyum bitki dokularının içinde ve üzerinde büyüdükçe sklerotia oluşacaktır. Bu sklerotiler sezon boyunca daha fazla apothecia oluşturmak için çimlenmeyecek, bunun yerine hasat ve toprak işleme işlemleri sırasında toprağa geri dönecek ve gelecekteki duyarlı bir ürün için inokülüm (enfektif mantar kaynağı) olmak üzere kışı geçirecektir. Sclerotia oldukça dayanıklı yapılardır ve toprakta uzun süre hayatta kalabilirler.
Kaynak: http://www.ndsu.edu/pubweb/~bernelso/soydiseases/sclerotinia.shtml
FieldClimate'de Sclerotinia için model yağmurlu dönemlere, bağıl neme ve sıcaklığa ve yaprak ıslaklığına bağlı olarak hesaplanır. Uzun yağışlı dönemlerde fungal patojen tarafından bir appressorium oluşturulması yoluyla bir enfeksiyon önerilmektedir. Diğer bir enfeksiyon yolu ise "hidrolitik enfeksiyon" olarak adlandırılır. Bu yol, bitki kütikulasını, orta lamelleri ve birincil ve ikincil hücre duvarlarını ve bitkinin tamamını sırayla bozan hidrolitik enzimlerin salınmasına dayanır. Ayrıntılı olarak açıklanan model bu sayfada.