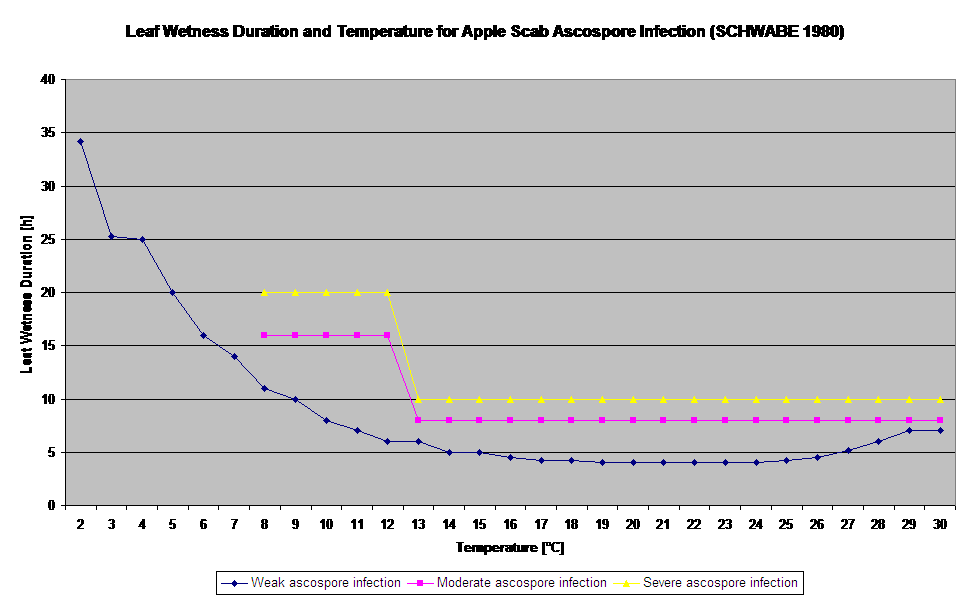
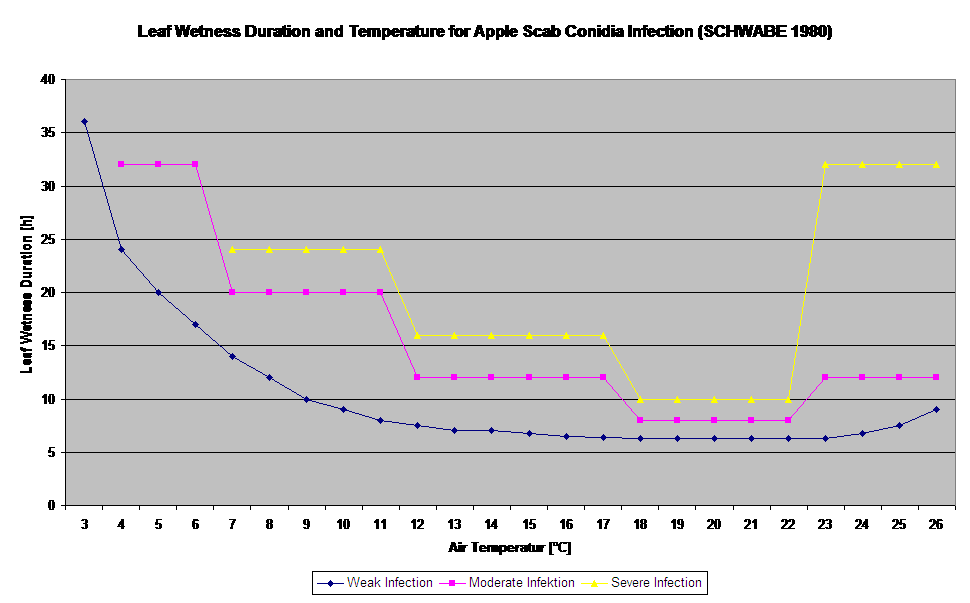
Germinația și pătrunderea ascosporilor de Venturia inaequalis pe frunze sau fructe depind de perioadele de umezeală a frunzelor (frunze umede sau umiditate relativă >80% (menține nivelul de infecție până sub 80% r.h.). și de temperatura aerului (peste 2°C). Temperaturile mai ridicate conduc, în condiții de umezeală, la o pătrundere mai rapidă, în timp ce la temperaturi mai scăzute sunt necesare perioade mai lungi de umezeală a frunzelor. Această relație a fost publicată pentru prima dată de MILLS și LAPLANTE (1945). Calculul nostru este efectuat pe baza publicațiilor lui SCHWABE (1980). SCHWABE a descris severitatea infecțiilor de tărtăcuță în funcție de temperatură. Aceste observații sunt în concordanță cu propriile noastre observații.
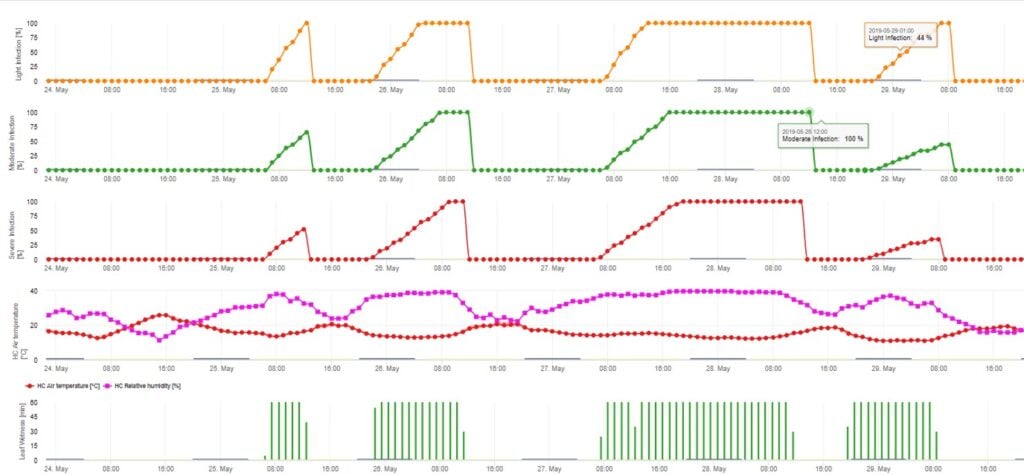
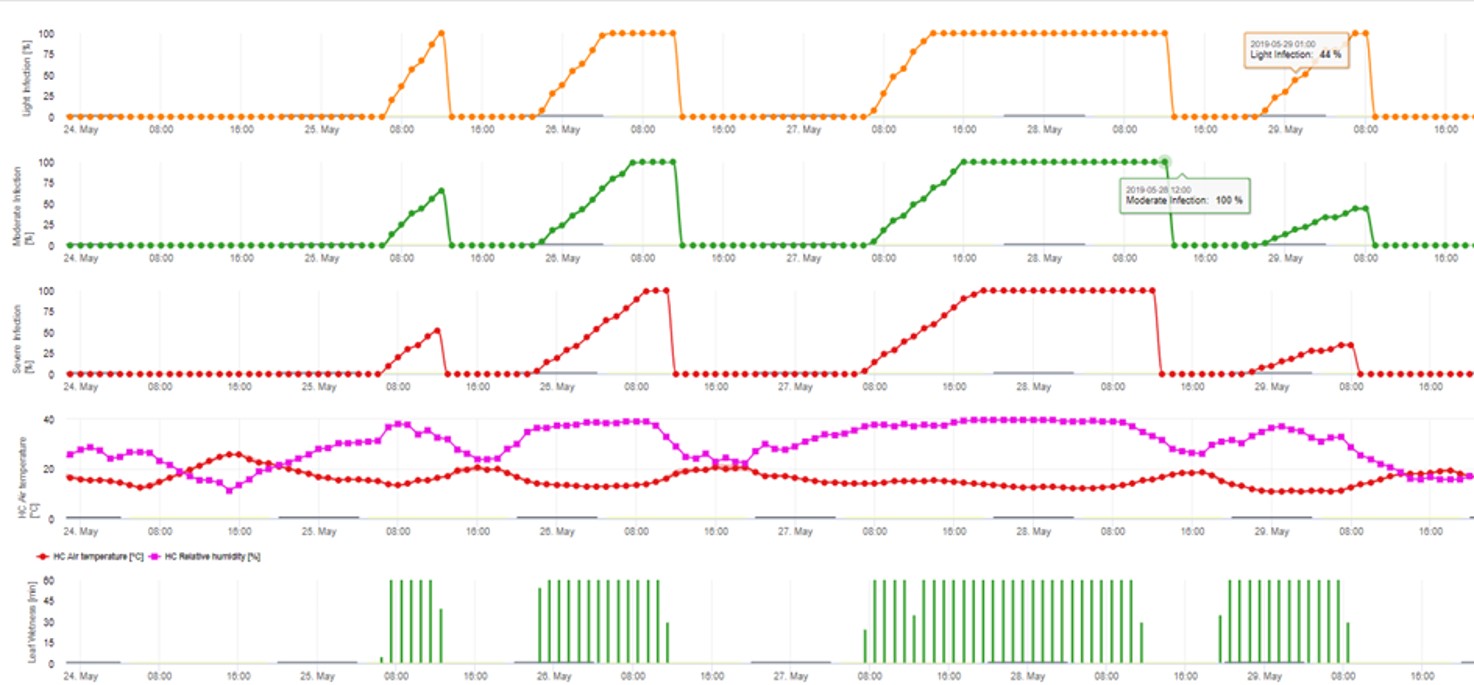
Modelul de infecție cu ascospori calculează evoluția infecției, în funcție de umiditatea și temperatura frunzelor, pentru infecții slabe, moderate și severe. Împărțirea predicției în trei "clase de gravitate" este foarte tradițională pentru modelele de scorbură a mărului. Progresul infecției este afișat sub formă de grafic pentru valorile orare. O infecție a avut deja loc de îndată ce curba de progres pentru o infecție slabă atinge 100%. O infecție moderată sau severă este completă dacă curbele de progres pentru aceste clase de gravitate ating (100%). Adesea, în practică sau în realitate, o infecție ajunge la 70% sau 80% dimineața devreme și, în funcție de condițiile meteorologice actuale, veți putea ști dacă această infecție se va termina sau nu. Afișarea valorilor maxime zilnice ale curbei procesului de infectare oferă o imagine de ansamblu asupra datelor de infectare preconizate în timpul sezonului.
Utilizarea primară a modelului
Pentru piață, este absolut important să avem fructe de măr fără boli pentru a obține un preț bun. Prin urmare, cultivatorii au tendința de a nu avea deloc toleranță la tărtăcuță în livezile lor. Modelul ar trebui să fie implementat într-un program de aplicare curativă (aplicarea de stropiri care conțin compuși de ciprodinil sau pirimetanil înainte de înflorire) atunci când modelul a indicat o infecție (indiferent dacă este slabă sau severă). În perioada de înflorire și la începutul perioadei de după înflorire, infecțiile severe sunt cele mai frecvente, iar toate perioadele umede au fost acoperite cu stropiri de protecție. În continuare, țesutul plantelor crește foarte repede, de aceea o protecție poate funcționa doar timp de 4 zile. În acest caz, ar putea fi necesară și o pulverizare curativă după o perioadă de infecție incompletă. În acest caz, însă, este important dacă infecția a fost slabă sau severă și dacă numărul de ascospori pregătiți pentru infecție a fost mare sau mic.
Literatură
- Schwabe WFS, 1980. Cerințe de umezeală și temperatură pentru infectarea frunzelor de măr de către Venturia inaequalis în Africa de Sud. Phytophylactica, 69-0.
- Schwabe WFS, 1980. Condițiile meteorologice care favorizează infecția cu tărtăcuță a mărului în Africa de Sud. Phytophylactica 12, 213-217.
- Mac Hardy W.E., 1996. Biologia, epidemiologia și managementul manei. St. Paul, MN: Am. Phytopathol. Soc.: 545.
Infecții secundare prin conidii
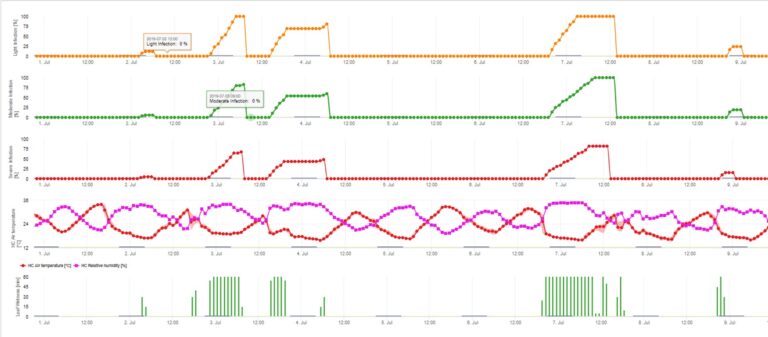
Infecțiile cu conidii (asexuate) depind de aceiași factori ca și infecțiile cauzate de ascospori. Diferența constă în faptul că conidiile pot fi evacuate în condiții de lumină și de întuneric, iar infecțiile conidiene au loc mai ales în timpul verii.
În funcție de regimul temperaturii aerului, conidiile de Venturia inaequalis au nevoie de perioade mai lungi sau mai scurte de umezeală a frunzelor pentru a germina și a pătrunde în frunzele sau fructele mărului. Această relație a fost publicată pentru prima dată de MILLS și LAPLANTE (1945). Calculele noastre sunt efectuate pe baza publicațiilor lui SCHWABE (1980). În timp ce Mills și LAPLANTE (1945) au presupus doar că conidiile au nevoie de o anumită perioadă de timp de umezeală a frunzelor; SCHWABE (1980) a inclus, de asemenea, importanța temperaturii în procesul de infectare.
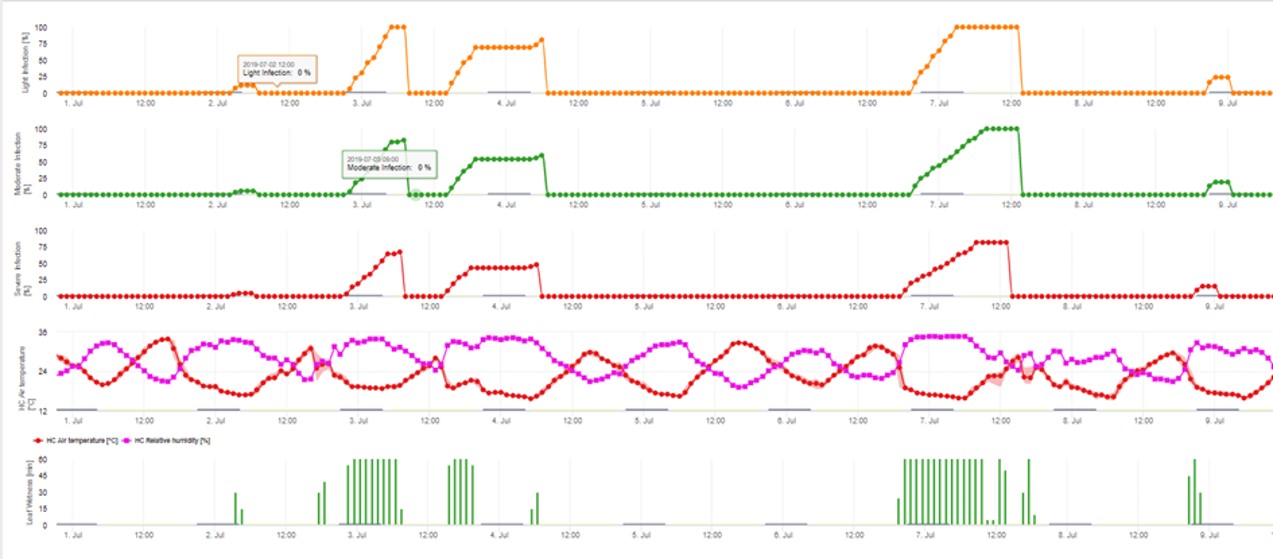
Modelul de infectare a conidiilor calculează evoluția infecției în funcție de umiditatea și temperatura frunzelor pentru infecții slabe, moderate și severe. Împărțirea predicției în "trei clase de gravitate" este foarte tradițională pentru modelele de combatere a tărtăcuței mărului. Progresul infecției este afișat sub formă de grafic pentru valorile orare. O infecție a avut deja loc de îndată ce curba de progres pentru o infecție slabă atinge 100%. O infecție moderată sau severă este completă dacă curbele de progres pentru aceste clase de gravitate ating 100%. Adesea, în practică sau în realitate, o infecție atinge 70% sau 80% dimineața devreme și, în combinație cu vremea reală, veți putea ști dacă această infecție se va termina sau nu. Afișarea valorilor maxime zilnice ale curbei de evoluție a infecției oferă o imagine de ansamblu asupra datelor de infecție preconizate în timpul sezonului.
Literatură:
- Infecția cu Venturia inaequalis, indicând datele de infecție cu ascospori și conidii (SCHWABE, W. 1980)
- Mac Hardy W.E., 1996. Biologia, epidemiologia și managementul manei. St. Paul, MN: Am. Phytopathol. Soc.: 545.
Prezentarea grafică a modelului
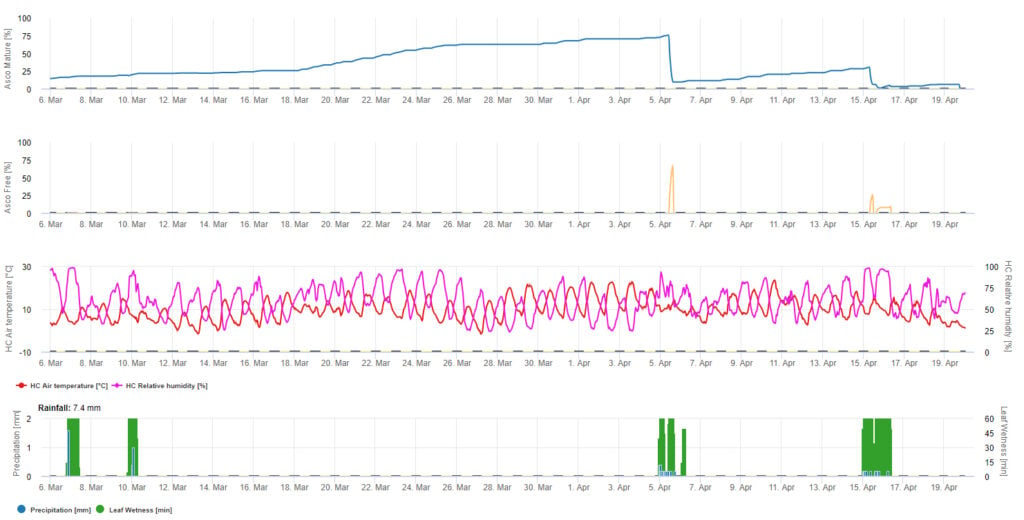
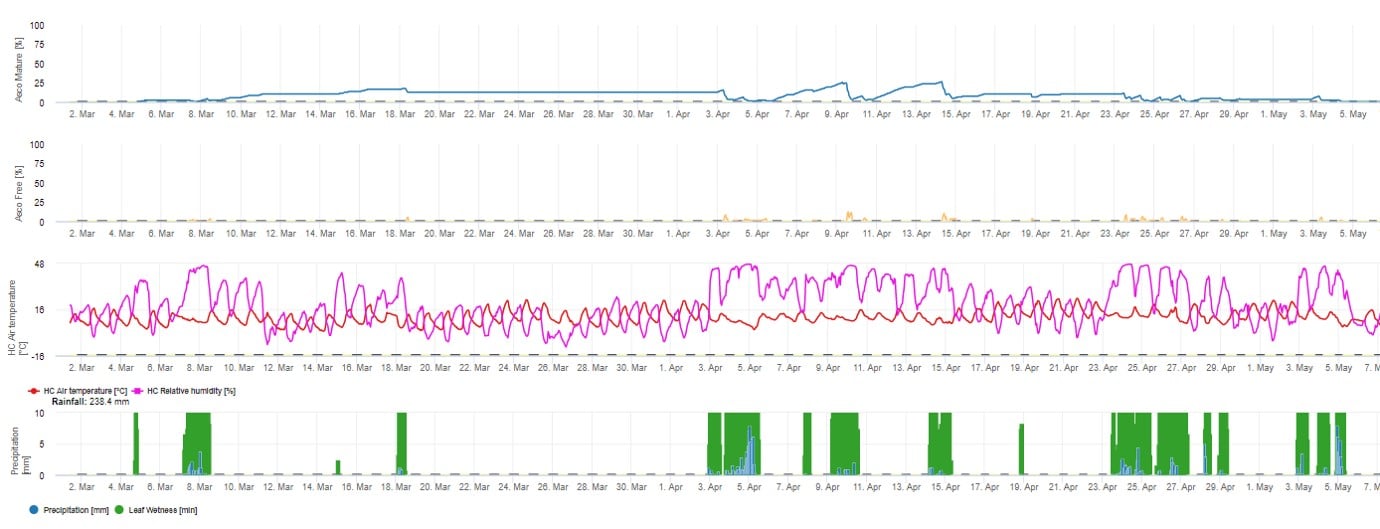
Pentru prezentarea grafică a modelului de maturare, formare și descărcare a ascosporilor se utilizează valori zilnice. Acesta arată că perioadele de umezeală a frunzelor conduc la eliberarea de ascospori și curbe pentru ascospori maturi și eliberați. Gravitatea eliberării și numărul de ascospori maturi nu pot fi evaluate în valori absolute. Valorile cuprinse între 0 și 100% indică cantitatea relativă de ascospori maturi și eliberați în funcție de condițiile climatice.
1. Maturizarea, formarea și evacuarea ascosporilor:
2. Infestarea cu ascospori se prezintă în trei clase diferite de gravitate (slabă: portocaliu, moderată: verde, severă: roșu). Ori de câte ori s-a ajuns la infecția 100%, s-au determinat condițiile optime pentru ca ascosporul să infecteze țesutul vegetal. Prin urmare, trebuie luate în considerare măsurătorile de protecție a plantelor (înainte de infecție, profilactic, după infecția 100%, curativ):
3. Infestarea cu conidii se prezintă în trei clase diferite de gravitate (slabă: portocaliu, moderată: verde, severă: roșu). Ori de câte ori s-a ajuns la infecția 100%, s-au determinat condițiile optime pentru ca ascosporul să infecteze țesutul vegetal. Prin urmare, trebuie luate în considerare măsurătorile de protecție a plantelor (înainte de infecție, profilactic, după infecția 100%, curativ):
Merele cultivate în scop comercial trebuie să fie lipsite de păduche. Merele cu crustă vor fi vândute numai pentru prelucrare. Prin urmare, scopul tuturor activităților de protecție a plantelor, atât în cultura convențională, cât și în cea ecologică, este de a avea fructe fără păduchi. Modelele, care arată descărcarea ascosporilor și infecția ascosporilor/conidiei sunt instrumente foarte importante pentru atingerea acestui obiectiv.
În sistemele de cultură convenționale se folosesc două tipuri de bază de fungicide împotriva manei: a) produse preventive, cum ar fi Captan, Mancozeb, Dithianone și Strobilurins sau b) produse curative, cum ar fi Cyprodinil (Chorus) sau Pyrimethanil (Scala) sau, pentru aplicarea în perioadele mai calde ale sezonului, fungicidele DMI. De fapt, majoritatea cultivatorilor urmează o strategie preventivă. Cu toate acestea, o strategie preventivă practică nu este capabilă să protejeze complet pomii de măr, deoarece pomii de măr cresc și dezvoltă flori, fructe și frunze. Prin urmare, pulverizarea preventivă protejează doar pentru o perioadă de 4 până la 7 zile, în funcție de creșterea reală a pomului. Astfel de intervale de pulverizare înguste nu pot fi gestionate; prin urmare, cultivatorii vor integra experiența lor privind clima locală, prognoza meteo și modelele de combatere a manei în gestionarea pulverizării. Aceștia vor programa stropirile preventive pe baza experienței lor și a previziunilor meteorologice. Modelele de infecție cu tărtăcuță a mărului le vor arăta data exactă a infecției (slabă, moderată și severă), precum și modelul de descărcare a ascosporilor/conidiilor și, cu ajutorul experienței lor, vor putea estima importanța unei infecții. Acest lucru oferă posibilitatea de a acționa cu un produs curativ în cazul în care o infecție de păduche a mărului a fost prea mult timp după ultima pulverizare preventivă.
În producția ecologică de mere, sulf de var s-a dovedit a fi cel mai eficient agent de control împotriva tărtăcuței. Controlul optim poate fi obținut dacă este pulverizat cu puțin timp înainte de infecție sau la începutul infecției. Acest lucru trebuie planificat în funcție de prognoza meteo. Uneori vom rata această perioadă optimă și va trebui să pulverizăm pe frunzele umede ale unei infecții aproape complete de păduche. Acest lucru va da totuși o eficacitate bună. Modelele privind păduchele de măr ne ajută să decidem dacă este nevoie de o pulverizare de emergență într-o infecție aproape completă.
Simptomele focului bacterian
Cankerii care iernează, care adăpostesc agentul patogen al focului bacterian, sunt adesea vizibili în mod clar pe trunchiuri și pe ramurile mari ca zone de coajă decolorată, de la ușor până la adânc deprimată, care sunt uneori crăpate pe margini. Cu toate acestea, cel mai mare număr de coșuri sunt mult mai mici și nu se disting atât de ușor. Aceștia apar pe ramurile mici unde au apărut infecții ale florilor sau lăstarilor în anul precedent și adesea în jurul tăieturilor făcute pentru îndepărtarea ramurilor bolnave. Deoarece multe dintre aceste canistre se instalează mai târziu în timpul sezonului, ele nu sunt adesea puternic deprimate și rareori prezintă fisuri ale scoarței la margini. De asemenea, sunt adesea destul de mici, având o lungime mai mică de 2,5 cm, cu scoarță roșiatică sau purpurie, care poate fi acoperită cu mici corpuri de fructificație de ciuperci negre (mai ales Botryosphaeria obtusa, agentul patogen al putregaiului negru al mărului).
Simptomele de blossom blight apar cel mai adesea la una sau două săptămâni după înflorire și, de obicei, implică întregul ciorchine de flori, care se ofilește și moare, devenind maro la măr și destul de negru la pere. Atunci când vremea este favorabilă dezvoltării agentului patogen, pe flori pot fi observate globule de suspensie bacteriană. De asemenea, pintenul care poartă ciorchinii de flori moare, iar infecția se poate răspândi și distruge porțiuni din ramura de susținere. Vârfurile lăstarilor tineri infectați se ofilesc, formând un simptom foarte tipic de "cioban". Lăstarii mai bătrâni care se infectează după ce au fost dezvoltate aproximativ 20 de frunze pot să nu prezinte acest simptom de încovoiere la vârf. Pe măsură ce infecția se răspândește de-a lungul axei lăstarului, frunzele prezintă mai întâi dungi întunecate la mijlocul nervurilor, apoi se ofilesc și devin brune, rămânând strâns atașate de lăstar pe tot parcursul sezonului. La fel ca în cazul infecțiilor florilor, agentul patogen invadează și ucide adesea o porțiune din ramura care susține lăstarul infectat. Primul simptom la mugurii de apă și la lăstarii care sunt invadați sistemic de la cancerele active din apropiere este apariția unei decolorări galbene spre portocalii a vârfului lăstarului înainte de ofilire. În plus, pețiolurile și nervurile medii ale frunzelor bazale ale acestor lăstari devin de obicei necrotice înaintea celor de la vârful lăstarului.
În funcție de soi și de stadiul de dezvoltare al acestuia în momentul în care se produce infecția, o singură floare sau un singur lăstar infectat poate duce la moartea unui membru întreg, iar în cazul în care este invadată frunza centrală sau trunchiul copacului, o mare parte din copac poate fi distrusă într-un singur sezon. În general, infecțiile de orice tip care apar între căderea petalelor și formarea mugurilor terminali duc, de obicei, la cele mai mari pierderi de ramuri și de arbori. În plus, arborii puternic structurați tind să sufere pierderi mai puțin severe de ramuri decât cei antrenați în sisteme mai slabe pentru o productivitate ridicată. În cazul în care portaltoii de măr foarte sensibili (M.26, M.9) sunt infectați, o mare parte a trunchiului și a ramurilor principale de deasupra altoiului rămân, de obicei, fără simptome, în timp ce în jurul portaltoiului se dezvoltă o caniculă maro închis distinctă. Pe măsură ce acest cancer al portaltoiului înconjoară pomul, partea superioară prezintă simptome de declin general (culoare slabă a frunzelor, creștere slabă) până la jumătatea sau la sfârșitul sezonului. În unele cazuri, frunzișul pomilor afectați de arsura portaltoiului dezvoltă o culoare roșie de toamnă timpurie la sfârșitul lunii august până la începutul lunii septembrie, nu foarte diferită de cea asociată adesea cu boala putregaiului de guler cauzată de o ciupercă din sol. Este posibil ca unii arbori cu infecții ale portaltoiului să nu prezinte simptome de declin până în primăvara următoare, moment în care pot fi observate canicule care se extind în sus în partea inferioară a trunchiului.
Ciclul bolii
Agentul patogen bacterian care cauzează arsura bacteriană iernează aproape exclusiv în caniculă pe ramurile infectate în sezonul precedent. Cel mai mare număr de coșuri și, prin urmare, cele mai importante pentru aportul de inoculum, apar pe ramurile cu diametrul mai mic de 38 mm, în special în jurul tăierilor efectuate în anul precedent pentru îndepărtarea ramurilor afectate de buboaie. La începutul primăverii, ca răspuns la temperaturile mai ridicate și la dezvoltarea rapidă a mugurilor, bacteriile de la marginile canceroșilor încep să se înmulțească rapid și produc un exudat gros, de culoare gălbuie până la albă, care se elaborează pe suprafața scoarței până cu câteva săptămâni înainte de perioada de înflorire. Multe specii de insecte (în special muștele) sunt atrase de acest exudat și, ulterior, dispersează bacteriile în toată livada. Odată ce primele câteva flori deschise sunt colonizate de bacterii, insectele polenizatoare transferă rapid agentul patogen la alte flori, ceea ce determină apariția altor cazuri de blossom blight. Aceste flori colonizate sunt supuse infecției în câteva minute după orice eveniment de umezire cauzat de ploaie sau rouă puternică, atunci când temperaturile medii zilnice sunt egale sau mai mari de 16 °C, în timp ce petalele florilor sunt intacte (receptaculii florilor și fructele tinere sunt rezistente după căderea petalelor). Odată ce apar infecțiile florilor, se pot aștepta simptome timpurii odată cu acumularea a cel puțin 57 de grade-zi (DD) mai mari de 13 °C, ceea ce, în funcție de temperaturile zilnice, poate necesita între 5 și 30 de zile calendaristice.
Odată cu apariția simptomelor de blossom blight, numărul și distribuția surselor de inoculum în livadă cresc foarte mult. Inoculul din aceste surse este răspândit în continuare de vânt, de ploaie și de multe insecte care vizitează ocazional vârful lăstarilor tineri, ceea ce crește probabilitatea apariției unui focar de arsură a lăstarilor. Cercetări recente efectuate în Pennsylvania indică faptul că hrănirea afidelor nu contribuie la apariția arsurii lăstarilor. Sunt necesare mai multe cercetări pentru a determina dacă șopârlele joacă sau nu un rol în incidența arsurii lăstarilor. Cele mai multe infecții ale vârfului lăstarilor se produc între momentul în care lăstarii au aproximativ nouă-zece frunze și apariția mugurilor terminali, atunci când sunt disponibile surse de inoculum și insecte vectori, iar temperaturile zilnice au o medie de 16 °C sau mai mult.
În anii în care nu se produc infecții ale florilor, sursele principale de inocul pentru faza de ofilire a lăstarilor sunt canistrele care iernează și, în special, lăstarii tineri de apă din apropierea acestor canistre, care se infectează pe măsură ce bacteriile pătrund în ei în mod sistemic de la marginile canistrelor. Astfel de infecții sistemice ale lăstarilor, denumite "canker blight", se pare că sunt inițiate la aproximativ 111 DD mai mari de 13 °C după vârful verde, deși este posibil ca simptomele vizibile să nu apară decât după acumularea a cel puțin 157 DD mai mari de 13 °C după vârful verde. În absența infecțiilor din flori, dezvoltarea infecțiilor de arsură a lăstarilor este deseori localizată în jurul zonelor în care se află cancerozități care au iernat.
Bășicarea florilor de model
Modelul cere utilizatorului să recunoască evenimentele locale specifice și în continuă schimbare și aspectele livezii sale care pot crește sau scădea riscul de arsură bacteriană în raport cu alte livezi din regiune. Modelul presupune ca utilizatorul să presupună că există un risc de infectare cu focul bacterian ori de câte ori florile sunt prezente pe pomi, în special în perioada de cădere a petalelor și în perioada de "postflorire", când pot rămâne flori împrăștiate pe multe soiuri de mere și pere. Utilizatorul modelului este rugat să evalueze cu atenție situația din locația sa specifică și să inițieze măsuri de control în cazul în care florile sunt prezente, nivelurile de risc sunt "ridicat" sau "extrem" și este probabil ca umezirea florilor să aibă loc cândva în următoarele 24 de ore.
Structura modelului: Temperaturi și umezeală: Principalul proces de foc bacterian care trebuie modelat este potențialul de dezvoltare bacteriană pe stigmatul florilor de măr și de pară. Această creștere depinde de temperatură, astfel încât o predicție fiabilă a riscului de infectare necesită utilizarea unei metode de măsurare care să reflecte cu cea mai mare precizie creșterea bacteriilor. Erwinia amylovora colonii. Principalul dezacord între modelatori este modul în care ar trebui să se facă acest lucru.
Modelul Cougar blight estimează rata de creștere bacteriană în funcție de gradele-oră pe baza unei curbe specifice a ratei de creștere. Această curbă de creștere se bazează pe rata de creștere a E. amylovora bacterii în testele de laborator. Valorile gradelor-oră sunt acumulate pentru fiecare oră din zi în care temperaturile depășesc 15 °C. Valorile orare cresc pe măsură ce temperaturile cresc de la 15 °C la 29 °C, scad la temperaturi mai ridicate și ajung la zero pentru orice oră cu temperaturi de peste 40 °C.
În FieldClimate.com, ambele modele sunt afișate în aceeași figură. Modelul Fireblight DIV se bazează pe Courgarblight, iar modelul Blossomblight se bazează pe Maryblight.
Model cougar blight
Incidența arsurii florilor este unul dintre cele mai sporadice aspecte ale epidemiilor de arsură bacteriană. Modelul a fost construit pornind de la ipoteza că există o abundență de inocul și că, pentru ca un eveniment de infectare a florilor să se producă, trebuie să fie îndeplinite în succesiune patru condiții stricte. Aceste condiții sunt:
1) floarea trebuie să fie deschisă cu stigmatele și petalele intacte, stigmatele trebuie să fie expuse pentru colonizare, florile în căderea petalelor sunt rezistente;
2) acumularea a cel puțin 110 °C ore > 18,3°C în ultimele 44°C zile > 4,4°C definește potențialul de infecție epifită pentru cea mai veche floare deschisă și, prin urmare, cea mai colonizată floare din livadă.
3) un eveniment de umezire care se produce sub formă de rouă sau 0,2 mm de ploaie sau 2,5 mm de ploaie în ziua precedentă permite deplasarea bacteriilor de la stigmatul colonizat la nectardoze.
4) o temperatură medie zilnică de >= 15,6°C: Acest lucru poate influența rata de migrare a bacteriilor în nectarthode, precum și multiplicarea bacteriilor necesare pentru a stabili infecții.
Prezentarea grafică a modelelor
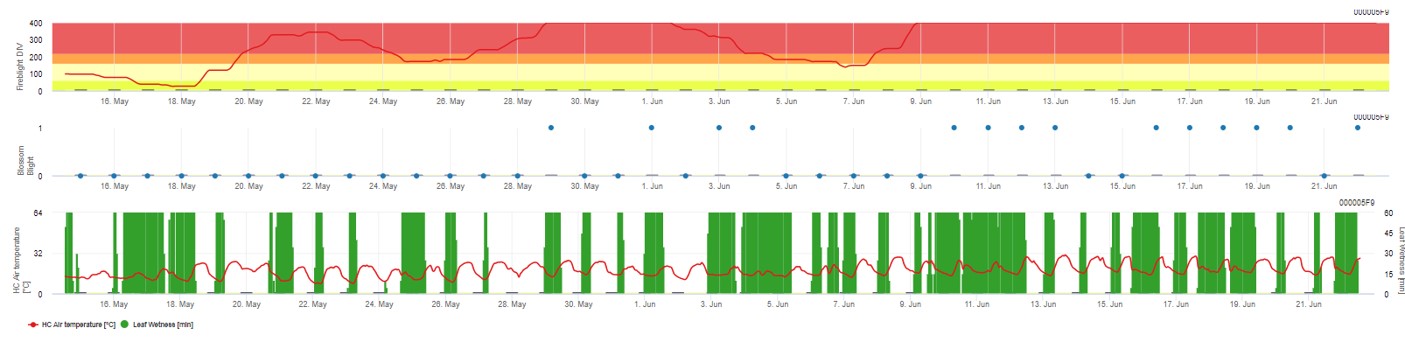
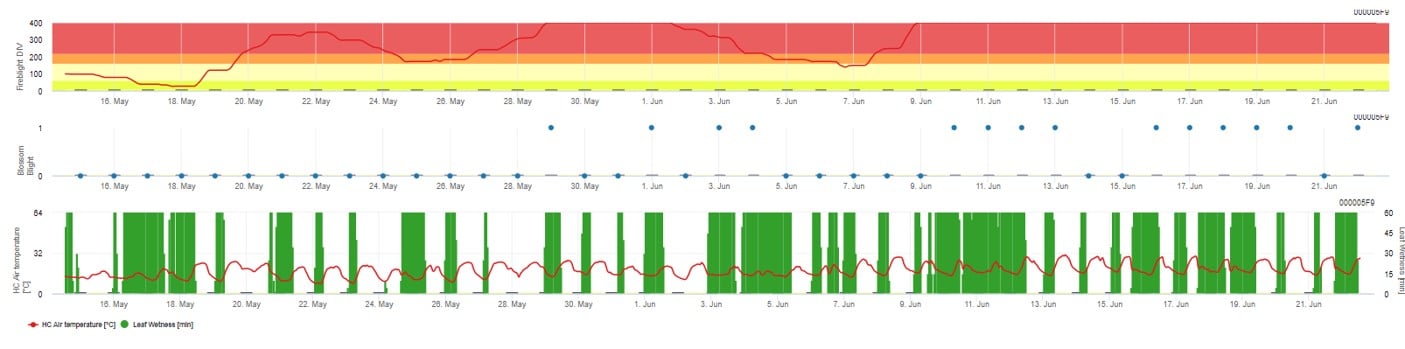
Ambele modele sunt afișate pe aceeași pagină. Prezentarea grafică a modelelor Fire Blight este cel mai bine utilizată cu valori agregate pe bază zilnică. Graficul prezintă valorile climatice legate de această boală. Rezultatul Cougar Blight numit Fire Blight DIV și rezultatele Blossom Blight. Pentru a interpreta rezultatele Cougar Blight, graficul este subînțeles de 5 culori diferite. Distribuția acestor culori se face pe baza setărilor privind istoricul primului buboi al livezii (a se vedea butonul din partea dreaptă sus). Cele 5 culori indică clasa de risc pentru valorile DIV.
Interpretarea valorilor (în funcție de apariția incendiilor din ultimii ani, configurația de bază):
Prezent în apropiere de caniculă activă
Setați setările pentru istoricul focului bacterian cu "Settings" (Setări) în partea superioară dreaptă:
Afișarea rezultatelor modelului Fire Blight prin 1) Modell Cougar Blight (numit Fireblight DIV pe baza istoricului și a culorilor de fundal) și 2) Modell Blossom Blight
| Potențialul de prezență a agenților patogeni | Scăzut | Moderat | Mare | Extreme |
| Potențialul de prezență a agenților patogeni | Scăzut | Moderat | Mare | Extreme |
| Fără foc bacterian în ultimii doi ani | 0-350 | 350-500 | 500-800 | 800+ |
| Focul bacterian în zona locală două sezoane | 0-300 | 300-500 | 500-750 | 750+ |
| Focul bacterian în zona locală două sezoane | 0-250 | 250-450 | 450-700 | 700+ |
| Focul bacterian din livada de anul trecut | 0-200 | 200-350 | 350-500 | 500+ |
Prag de acțiune:
Se recomandă controlul în cazul în care se detectează o perioadă de infecție cu risc ridicat sau extrem. Factorii care determină pagubele sunt: numărul de flori, vârsta pomului, vigoarea și soiul,... de asemenea.
Posibilitatea unei infecții cu Blossom Blight este afișată printr-o bară de la 0 la 1 (deci toate condițiile sunt îndeplinite sau nu).
Scopul modelelor de combatere a focului bacterian este de a evalua probabilitatea unei infecții prin Erwinia amyloflora în livadă.
Modelul Mary Blight, care evaluează pentru blossom Blight, indică foarte bine situațiile de infecție cu impact economic ridicat. Din acest motiv, este utilizat destul de frecvent pentru a indica utilizarea antibioticelor împotriva acestui agent patogen. Cougar blight oferă informații despre riscul de infecții cu focul bacterian din cauza posibilităților generale de propagare a agentului patogen. Ponderarea sa în funcție de istoricul unei livezi este foarte utilă pentru a ne indica cât de atent trebuie să verificăm livezile pentru a depista simptome de arsură bacteriană, chiar și în situațiile în care Mary Blight nu indică o infecție.
Biologia moliei Codling
The C. pomonella își petrece iarna sub formă de larvă matură, înfășurată într-un cocon care se află în zonele protejate ale copacului (crăpături sau fisuri ale scoarței), care este locul preferat, sau la baza copacului, în litiera de la sol. În livezile moderne de mere cu densitate mare, există foarte puține locuri de cocoonare pe copaci din cauza scoarței netede. Pupele se formează în coconul țesut de larvele mature.
Stadiul adult iese din carcasa pupilară și, după o scurtă perioadă de întărire a exoscheletului, zboară în copac. Cea mai mare parte a literaturii de specialitate indică faptul că activitatea moliilor are loc în treimea superioară a coronamentului copacilor. Adulții sunt activi la asfințit timp de câteva ore, cu un prag mediu de activitate cuprins între 13°C și 15°C. Împerecherea are loc, de obicei, pe gazdă, iar depunerea ouălor are loc imediat după aceea. În serile călduroase, este posibil ca o femelă de molie să iasă, să se împerecheze și să depună ouă într-o perioadă de două zile. A C. pomonella molia poate trăi mai mult de 20 de zile în condiții ideale, însă în câmp este probabil ca aceasta să supraviețuiască mult mai puțin timp, mai ales în timpul verii fierbinți. În condiții de laborator, o femelă de molie depune până la 100 de ouă, dar fecunditatea reală în câmp este probabil mai mică și majoritatea sunt depuse în primele câteva zile de la apariție și împerechere.
Ouăle sunt depuse individual pe suprafața superioară a frunzei, pe crenguțe sau pe fructe. La prima generație, ouăle sunt depuse numai pe frunze până la aproximativ două săptămâni după ce fructele au prins contur, atunci când se pierde puful de pe fructe. La a doua generație, aproximativ 65% din ouă sunt depuse pe frunze, dar acestea sunt de obicei foarte aproape de un fruct. Ouăle sunt de culoare alb-crem când sunt depuse pentru prima dată, au un inel roșu la jumătatea dezvoltării, iar când sunt aproape de ecloziune se poate vedea capsula neagră a capului.
O larvă abia eclozată rătăcește până când găsește un fruct. Ea mestecă prin piele și se hrănește cu pulpa mărului de sub piele timp de câteva zile. În procesul de mestecare a pielii și de hrănire dedesubt, larvele împing zgura în afara zonei de intrare. Adesea există un inel roșu care înconjoară intrarea, în special în cazul fructelor tinere. După ce larva trece la stadiul următor, aceasta găurește spre miez, unde se va hrăni cu semințe până la maturitate. Când este aproape matură, larva face un tunel de la miez spre exteriorul mărului și astupă gaura cu excremente și mătase. Când este pregătită pentru a înfășura un cocon, larva părăsește fructul, plimbându-se prin copac sau căzând pe pământ pe un fir de mătase, și găsește un loc protejat.
Model pentru molia Codling
Modelul nostru se bazează pe acumularea temperaturilor efective între >= 10°C și <=32°C. În emisfera nordică, această acumulare începe cu data de 1 ianuarie. În emisfera sudică, aceasta începe la începutul lunii iulie. Acumulăm fiecare măsurătoare în echivalentul său pentru o zi (a se vedea graficul de mai jos). Pentru dezvoltarea insectelor se utilizează următoarele praguri:
*Generare completă= 650
| *De la depunerea ouălor până la eclozarea ouălor | = 88 |
| *Primul stadiu de larvă | = 60 |
| *Secunda etapă de larvă | = 65 |
| *Stadiul al treilea de larvă | = 55 |
| *Stadiul de larvă | = 45 |
| *Stadiul al cincilea de larvă și pupae | = 335 |
Graficul produs de model prezintă prima apariție și durata de apariție a larvelor de la stadiul 1 la 5 și prezența adulților în 5 niveluri.
| *Nivelul 1: | Activitate de zbor, fără ouat |
| *Nivelul 2: | Activitate de zbor cu posibila depunere de ouă. |
| *Nivelul 3: | Activitate de zbor cu depuneri de ouă. |
| *Nivelul 4: | Activitate de zbor cu depunere de ouă rezonabilă până la puternică. |
| *Nivelul 5: | Activitate de zbor cu depunere puternică de ouă. |
Prezentarea stadiilor adulte ale moliei Codling în 5 niveluri diferite (primul grafic), precum și a gradelor de zile acumulate (linia roșie, al doilea grafic) pentru dezvoltarea adulților și a stadiilor larvare prezentate în albastru (al treilea grafic).
Mai multe specii de afide sunt prezente în livezile de mere. Afidul verde al mărului, afidele mărului de ovăz sau afidele mărului și afidele rozaceelor. În grădinile private, mai ales, afidele rozalii au o anumită importanță. Acest afid duce la modificări hormonale puternice în vârfurile lăstarilor cu infestări. Frunzele se curbează și lăstarul se scurtează. Mai târziu, frunzele se îngălbenesc și mor. Acest afid afectează mai ales lăstarii tineri. Afidele rozacee de măr pot produce pagube grave deja într-o abundență mică. În timp ce afidele verzi ale mărului și afidele de mărul ierbos pot apărea în abundență mai mare, fără a duce la daune economice. Afidele de măr trebuie să fie considerate ca fiind de importanță neeconomică.
Afidele rozacee de măr
Aceasta este o specie europeană veche, introdusă în Statele Unite în jurul anului 1870. A devenit un dăunător major al merelor spre sfârșitul secolului al XIX-lea. Pentru a prospera, această specie trebuie să aibă o abundență de planta gazdă de vară, planta de plantare cu frunze înguste. Există un paralelism remarcabil între introducerea și răspândirea afidului roz al mărului și plantainul cu frunze înguste. Acest afid este prezent în toate zonele pomicole din întreaga lume.
Etapele vieții
Ouă: Oul este oval și ușor aplatizat pe partea dinspre scoarță. Lungimea variază între 0,49 și 0,56 mm. Când este depus pentru prima dată, este galben strălucitor și acoperit cu o substanță glutinică care se întărește cu vârsta. Culoarea se schimbă treptat în galben verzui și, în cele din urmă, într-un negru de jet strălucitor. Timpul necesar pentru această schimbare de culoare variază, în condiții normale de exterior, de la aproximativ nouă zile la peste două săptămâni.
Nimfă: Toți indivizii care eclozează din ouă sunt femele viviparoide fără aripi. Există cinci stadii. Ultimul este mama tulpină matură, care, la scurt timp după cel de-al patrulea stadiu, începe să producă pui vii pe cale partenogenetică. Ea produce în medie cinci-șase pui pe zi. Nimfele din a doua generație, care sunt toate femele, ajung la maturitate în două-trei săptămâni. Marea majoritate a nimfelor încep să se reproducă pe măr, deși câteva dintre ele pot dezvolta aripi și pot migra la plantain.
A treia generație se produce în iunie și la începutul lunii iulie. Majoritatea acestei generații își dezvoltă aripile și migrează către plantain cu frunze înguste. În unele anotimpuri, femelele fără aripi din a treia generație produc o a patra generație pe măr. În ultimii ani, s-a observat în unele zone că populațiile dăunătoare de afide rozalii ale mărului au persistat în livezi până la mijlocul verii sau mai târziu. Nu se știe dacă această schimbare se datorează procesului de selecție, schimbărilor de obicei sau lipsei de nevoie de o gazdă alternativă.
Adulți: Adultul variază considerabil în ceea ce privește marcajele de culoare. Culoarea generală este maro-roz, cu o nuanță rozalie datorată unui înveliș pulverulent. Unii dintre adulții mai în vârstă sunt purpurii, în timp ce adulții mai tineri sunt în mod hotărât roz-roșiatic.
Gama de gazde
Mărul este gazda preferată, dar afidele se hrănesc și cu pere și păducel. Cortland, Golden Delicious, Rhode Island Greening și Ida Red sunt soiuri de mere deosebit de sensibile. Afidul se găsește în toate zonele pomicole din Statele Unite și Canada.
Rănire sau deteriorare: Hrănirea cu afidele rozacee ale mărului provoacă adesea curbarea frunzelor de măr, începând cu căderea petalelor. Aceste frunze pot deveni mai târziu de un roșu aprins. Hrănirea frunzelor din jurul ciorchinilor de fructe duce adesea la ciorchinarea, atrofierea și malformația fructelor în curs de dezvoltare. Aceste anomalii se agravează pe măsură ce fructele se dezvoltă și, în cele din urmă, pot face ca fructele să nu mai poată fi comercializate.
Populațiile mari de afide pot produce cantități mari de mierea de albine ca deșeu din seva cu care se hrănesc. Mierea de miere excretată pe fructe va servi drept teren de creștere pentru ciuperca mucegaiului de funingine, care va afecta finisajul mărului. Toxinele din saliva afidelor servesc, de asemenea, drept "picătură de oprire", împiedicând absciziunea (separarea naturală de pom) fructelor la recoltarea normală.
Istoria vieții: Afidele trec iarna în stadiul de ou. Ecloziunea are loc la începutul primăverii, cu aproximativ o săptămână până la 10 zile mai târziu decât în cazul afidelor de la măr și aproximativ în același timp cu afidele de la măr. Ouăle eclozează atunci când mugurii încep să se deschidă primăvara, pe o perioadă de două săptămâni. Imediat ce eclozează, puii caută mugurii de măr care se deschid; se pare că preferă mugurii de fruct. Ei se hrănesc pe partea exterioară a mugurilor de frunze și a grupurilor de muguri de fructe până când frunzele încep să se desfășoare. Apoi își croiesc drum în interiorul ciorchinilor și încep să sugă seva din tulpini și din fructele nou formate.
Hrănirea lor face ca frunzele să se încovoaie, protejând afidele de spray-uri și de unii dușmani. Încolăcirea severă a frunzelor provocată de această specie este probabil cea mai caracteristică trăsătură a activității sale. O singură tulpină mamă situată pe partea inferioară a unei frunze, aproape de nervura mediană, va face ca frunza să se îndoaie strâns. Este nevoie doar de câteva mame de tulpină pentru a provoca o încovoiere severă a tuturor frunzelor din jurul unui mugure de floare care se deschide, oferind o protecție ideală afidelor care se dezvoltă rapid. Mamele de tulpină ajung la maturitate atunci când pomii de măr încep să înflorească.
Mamele de tulpină mature sunt foarte inactive. Ele se așează, se hrănesc și produc pui într-un ritm rapid. Atunci când sunt distribuite, își îndepărtează rapid ciocul de pe țesuturile plantelor și caută un alt loc în care să continue. Mamele de tulpină se maturizează la aproximativ două săptămâni după eclozare. Perioada de timp depinde în mare măsură de condițiile meteorologice. Producția de pui începe, de obicei, la două sau trei zile după ultima mutare și continuă fără întrerupere mai mult de o lună.
Producția totală a unei singure femele este în medie de aproximativ 185. În mod normal, perioada de reproducere se întinde de la începutul lunii mai până în iunie. De obicei, perioada maximă de activitate reproductivă este în jurul ultimei săptămâni din mai și primei săptămâni din iunie, când fructele tinere încep să se fixeze și încep să crească activ. Afidele rozacee ale mărului sunt rareori întâlnite atacând lăstarii tineri și în creștere rapidă. Acesta se limitează la frunziș, la tulpinile florale și la fructele tinere.
Una dintre trăsăturile caracteristice ale acestei specii este adunarea puilor în jurul mamei. Fiecare mamă tulpină individuală sau grup de mame va avea în jurul ei sute de pui. Frunzele infestate pot fi în curând acoperite - în unele cazuri, de mai multe straturi de afide. Acest obicei de a se aduna distruge în curând frunzele infestate și provoacă migrarea forțată a afidelor. Puii se deplasează activ și grăbit, părând nerăbdători să găsească un loc potrivit pentru a se hrăni. În această perioadă, ei sunt frecvent găsiți adunați pe fructele în formare sau atacând frunzele noi, suculente, care se desfășoară.
A doua generație a avut nevoie de patru până la 40 de zile pentru a ajunge la maturitate și a produce pui. Cea mai mare parte a celei de-a doua generații este formată din femele fără aripi. Producția medie totală a fiecărui individ din a doua generație este de aproximativ 119 exemplare. Obiceiurile și activitățile celei de-a treia generații nu diferă de cele ale celei de-a doua. Afidele se adună în număr mare pe partea inferioară a frunzelor, provocând o încovoiere puternică. Atacă, de asemenea, fructele încolțite și în curs de dezvoltare, producând leziuni caracteristice.
Cea mai mare parte a acestei a treia generații dobândește aripi și migrează către planta cu frunze înguste, planta gazdă de vară. Se poate produce o a patra generație. Toate acestea produc aripi și migrează către planta de plantare cu frunze înguste. După ultima mutație, adulții înaripați sunt foarte fragili și inactivi. Ei rămân secretați în frunzele încolăcite timp de două sau trei zile înainte de a se aventura în zborul lor migrator. Chiar înainte de zbor, devin foarte activi și nervoși, alergând de colo-colo sau mișcându-și aripile în sus și în jos în așteptarea zborului.
Pufarul verde al mărului
Afidul verde al mărului, Aphis pomi (DeGeer), este larg răspândit în toate zonele de cultivare a merelor. În emisfera nordică, acesta apare pentru prima dată în livezile de mere la sfârșitul lunii mai până la începutul lunii iunie. Insecta suge seva din frunzele de pe mugurii de apă și de pe creșterile terminale suculente. Afidele verzi ale mărului se găsesc, de obicei, în apropierea nervurilor principale, pe suprafața inferioară a frunzei. Afidele verzi ale mărului atacă, de asemenea, perele, păducelul, gutuile, mărul de crab și spiraea.
Descrierea afidei verzi a mărului
Ouăle afidei verzi a mărului sunt ovale și de culoare neagră lucioasă. Nimfele și adulții sunt de culoare verde închis sau verde deschis. Afidul verde al mărului are capul verde, abdomenul uniform verde sau verde-gălbui, picioarele negre și cornițele lungi și negre - "țevile cozii" (la capătul din spate al insectei). Femelele adulte sunt de culoare verde închis până la negru și au aripi. În timpul verii, aceste femele, sau alate, dau naștere la pui vii pe cale partenogenă.
Biologia afidei verzi a mărului
Afidul verde al mărului iernează sub formă de ouă pe drajoni, la baza mugurilor și pe cicatricile frunzelor din lăstarii terminali. Ouăle eclozează, de obicei, atunci când mugurii se sparg și primele frunze se desfășoară. Nimfele încep să se hrănească imediat cu frunzele în curs de dezvoltare. Inițial sunt prezente pe lăstarii terminali, iar mai târziu se mută pe frunzele mai bătrâne ale ciorchinelui. Afidele adulte femele produc femele fără aripi în două săptămâni, fiecare femelă fiind capabilă să producă 50-100 de urmași vii. Afidele tinere se dezvoltă în șapte până la zece zile. De obicei, populațiile de afide verzi ale mărului se formează pe măr încet la începutul primăverii (înflorire, petale, toamnă) și mai rapid pe măsură ce temperaturile medii zilnice cresc. Afidele înaripate, sau alate, și afidele fără aripi sunt produse în timpul verii. Afidele verzi ale mărului sunt cele mai numeroase în luna iulie și la începutul lunii august. Există mai multe nimfe decât adulți alați în stadiile inițiale ale unei infestări. Procentul de alate crește pe măsură ce populația crește. Coloniile de afide verzi ale mărului se pot dubla într-o săptămână dacă condițiile sunt favorabile. Dacă afidele nu au o creștere nouă suculentă și trebuie să se hrănească cu frunze mai bătrâne, atunci numărul de puiet produs poate scădea cu până la 50%. Dacă temperaturile zilnice sunt de 30-32°C și mai mari, femelele nu se reproduc bine. Atunci când temperaturile sunt ridicate timp de mai multe zile, afidele vor muri. În timpul verii se dezvoltă mai multe generații de afidele verzi ale mărului, iar formele înaripate se dispersează în toată livada. Uneori, afidul verde al mărului și afidul roz al mărului, Dysaphis plantaginea Pars., au colonii pe aceeași frunză. Afidul roz al mărului este roz sau purpuriu. Toamna, masculii și femelele fără aripi se dezvoltă și se împerechează, iar femelele depun ouă pentru iarnă. Cel mai mare număr de ouă se află la 15-20 cm de vârful crengilor. Ouăle sunt rareori pe ramurile mari ale schelelor sau pe trunchiurile pomilor de măr.
Daune
Afidele verzi ale mărului sug seva din frunze. Infestările puternice reduc vigoarea și creșterea lăstarilor. Acest lucru este deosebit de îngrijorător în pepiniere și în livezile tinere, care nu sunt pe rod. Insecta poate reduce dimensiunea mugurilor și lungimea internodală și poate provoca încovoierea frunzelor. Hrănirea afidelor poate stimula, de asemenea, creșterea ramurilor laterale care poate afecta forma pomilor. Încolăcirea frunzelor și terminalele slăbite sunt susceptibile la vătămări în timpul iernii. Mierea de miere produsă de afide se poate scurge pe fructe, permițând dezvoltarea ciupercilor de funingine. Ciuperca pătează fructele și scade valoarea de piață. Când infestările sunt puternice, afidele verzi ale mărului se pot hrăni cu mere imature și pot provoca ruginirea.
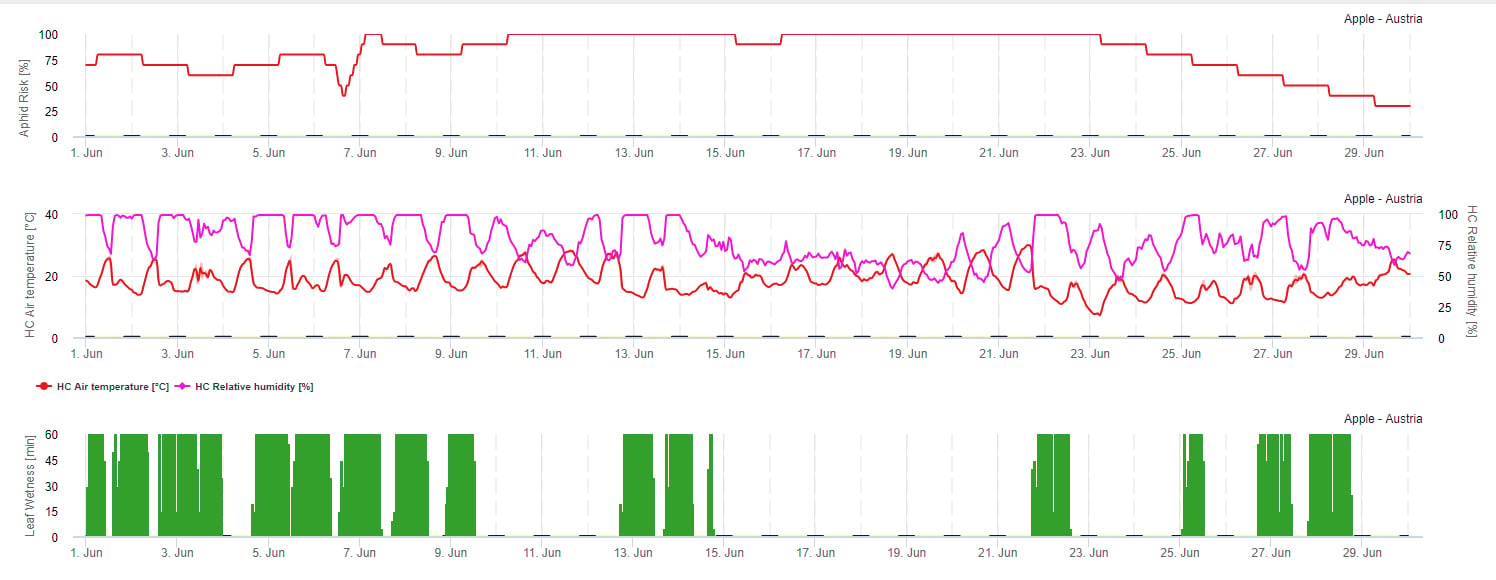
Modelul riscului de propagare
Înmulțirea afidelor este influențată în principal de temperatură. Cu cât mai multe ore sunt apropiate de temperatura optimă (20-32°C) și există o sursă de hrană disponibilă, cu atât mai mare va fi rata de înmulțire. În timp ce ouăle și instartele tinere sunt sensibile la aerul uscat. Umiditatea relativă scăzută poate afecta și ea populațiile de afide (sub 30% h.r.). În afară de insecticide, afidele sunt combătute de prădători și ciuperci parazite. Ciupercile parazite vor depinde de umiditate. Cele mai multe dintre aceste specii ca Beauvaria bassiana sunt favorizate de umiditatea liberă.
Riscul crește atunci când temperatura optimă este cuprinsă între 20 și 32°C și umiditatea relativă între 30 și 95% r. h. și scade atunci când este prea umed (umezeală foliară, ploaie sau umiditate relativă peste 95%), prea rece (temperaturi mai mici de 20°C) sau prea cald (temperaturi peste 32°C), nopți umede (umezeală foliară în orele de noapte).