La rouille du soja, en particulier la rouille asiatique du soja, est une maladie grave du soja causée par un champignon pathogène. Phakopsora pachyrhizi. Distribution de la maladie : Asie et Australie, et plus récemment d'Afrique, d'Amérique du Nord et d'Amérique du Sud. La maladie a été observée pour la première fois au Japon en 1902. L'agent pathogène a été distribué en Asie et en Australie par l'intermédiaire de l'Amérique. La rouille du soja ne peut pas passer l'hiver dans les régions où il fait froid, mais elle peut se propager par le vent sur de si grandes distances, son développement peut être si explosif et elle peut causer une perte si rapide des feuilles qu'elle est maintenant l'une des maladies les plus redoutées dans les régions productrices de soja du monde.
Symptômes et signes
Les premiers symptômes de la rouille du soja causée par le Phakopsora pachyrhizi commencent par de très petites taches brunes ou rouge brique sur les feuilles. Au champ, ces taches commencent généralement dans la partie inférieure de la canopée à la floraison ou après, bien que les semis puissent être infectés dans certaines circonstances. Souvent, les premières lésions apparaissent vers la base de la foliole, près du pétiole et des nervures de la feuille. Cette partie de la foliole retient probablement la rosée plus longtemps, ce qui rend les conditions plus favorables à l'infection. Les lésions restent petites (2-5 mm de diamètre), mais leur nombre augmente au fur et à mesure que la maladie progresse. Des pustules, appelées urédosores, se forment dans ces lésions, principalement sur la face inférieure des feuilles, et elles peuvent produire de nombreuses urédospores.
Les pustules en relief sont visibles à l'œil nu, surtout lorsqu'elles sont sporulées. Même si les lésions sont petites, chaque lésion comporte souvent plusieurs pustules (urédinia) . Les lésions peuvent être complètement recouvertes d'urédospores lorsque les pustules sont actives. Les urédinospores de la rouille du soja sont de couleur jaune-brun pâle à incolore, avec une ornementation de surface échinulée (épines courtes). Cette coloration est différente de celle de nombreux autres pathogènes de la rouille dont les spores sont souvent brun rougeâtre (couleur rouille). La germination de P. pachyrhizi Les urédospores se produisent à travers un pore équatorial (central), produisant un tube germinatif qui se termine par un appressorium, que le champignon utilise pour pénétrer dans l'hôte directement ou à travers un stomate.
Au fur et à mesure que des lésions se forment sur une foliole, la zone affectée commence à jaunir, et finalement la foliole tombe de la plante. Si la rouille du soja commence généralement dans la partie inférieure de la canopée, elle progresse rapidement vers le haut de la plante jusqu'à ce que toutes les feuilles présentent un certain niveau de maladie. Les plantes gravement malades peuvent être complètement défoliées. La perte de tissu foliaire efficace entraîne des réductions de rendement dues à des graines moins nombreuses et plus petites. Des pertes de rendement allant jusqu'à 30 à 80% ont été signalées, mais l'ampleur de la perte dépend du moment où la maladie commence et de la vitesse à laquelle elle progresse. Outre les feuilles, la rouille du soja peut également apparaître sur les pétioles, les tiges et même les cotylédons, mais la plupart des lésions de rouille se produisent sur les feuilles.
Les lésions peuvent être soit brunes, soit rouge-brun. Les lésions bronzées présentent de nombreuses pustules qui produisent de nombreuses urédospores. Les lésions rouge-brun, que l'on pense être une réaction de résistance modérée, ne présentent que quelques pustules qui ne produisent que quelques urédospores. Comme nous le verrons dans la section Gestion de la maladie, ce type de lésion dépend de la souche de l'agent pathogène, et peut apparaître sur la même feuille que les lésions bronzées, ou les lésions bronzées peuvent devenir rouge-brun avec l'âge. Les symptômes et les signes sur d'autres hôtes, comme le kudzu, sont similaires, bien que la taille des lésions puisse différer. En vieillissant, les pustules peuvent devenir noires. Ceci est dû à la formation d'une couche de téleutospores dans les pustules, transformant les pustules d'urédos en téleutospores.
Les téliospores ont deux fonctions : la survie du champignon en l'absence d'un hôte vivant (sursaison) et la reproduction sexuée. Les parois épaisses des téliospores protègent le champignon de l'environnement et des attaques d'autres organismes. Chez les rouilles, les téliospores germent en formant un basidium et quatre basidiospores au cours desquels se produit la recombinaison sexuelle. La germination des P. pachyrhizi Les téleutospores n'ont été observées qu'en laboratoire et ne semblent pas contribuer de manière significative à la perpétuation de cette maladie sur le terrain.
Biologie des agents pathogènes
Il existe deux champignons étroitement liés qui provoquent la rouille sur le soja : Phakopsora pachyrhizile pathogène de la rouille du soja, parfois désigné sous le nom de rouille asiatique ou australasienne, mais qui est désormais également présent dans l'hémisphère occidental, et P. meibomiaele pathogène de la rouille du soja dit du Nouveau Monde, qui ne se trouve que dans l'hémisphère occidental. À l'exception de quelques caractéristiques mineures, les deux champignons semblent morphologiquement identiques, mais... P. pachyrhizi est beaucoup plus agressif sur le soja que le P. meibomiae. A ce jour, P. meibomiae n'a pas été documenté comme causant des pertes de rendement significatives en Amérique centrale et du Sud. Les deux espèces peuvent être distinguées à l'aide de protocoles d'analyse de l'ADN.
Comme les autres rouilles, les agents pathogènes de la rouille du soja sont des parasites obligatoires qui ont besoin d'un hôte vivant pour se développer et se reproduire. Ils peuvent survivre loin de l'hôte sous forme d'urédospores pendant quelques jours seulement dans des conditions naturelles. Les deux agents pathogènes de la rouille du soja, à notre connaissance, ne produisent que deux types de spores : les urédospores et les téleutospores (figure 15). Cela contraste avec d'autres rouilles, qui peuvent avoir jusqu'à cinq stades de spores (par exemple, la rouille des tiges du blé). Pour la rouille du soja, comme pour la plupart des rouilles, le stade urédinien est le stade répétitif. Cela signifie que les urédospores peuvent infecter le même hôte sur lequel elles ont été produites (le soja) au cours de la même saison. Les épidémies peuvent se développer rapidement à partir de quelques pustules seulement, car les pustules productrices de spores sont produites en seulement 7 à 10 jours après l'infection, et chaque pustule peut produire des centaines d'urédospores. Des téliospores sont produites dans les anciennes lésions, mais elles ne semblent pas germer dans la nature, et on ne connaît pas d'hôte alternatif, ni d'écidies ou de spermogonies.
Sans la germination des téliospores, la reproduction sexuée ne peut avoir lieu. L'absence de reproduction sexuée devrait limiter la variabilité du champignon de la rouille, mais il existe néanmoins une variabilité substantielle dans P. pachyrhizi en ce qui concerne la virulence. Cela a limité l'utilisation de gènes uniques pour la résistance dans le soja, car en peu de temps, de nouveaux isolats de l'agent pathogène apparaissent et surmontent le gène de résistance. On ne sait pas comment cette variabilité trouve son origine dans les gènes de résistance. P. pachyrhizi. Rouille à bandes du blé, Puccinia striiformisa un cycle de vie similaire à celui de la P. pachyrhizi sans stade téleutosporien fonctionnel et donc sans reproduction sexuée, mais qui possède de nombreuses races. Il se peut que chaque gène de résistance soit si spécifique qu'une seule mutation dans le bon gène du champignon lui permet d'être virulent sur les hôtes possédant le nouveau gène de résistance.
Épidémiologie
Les épidémies de rouille du soja commencent avec l'arrivée de l'inoculum aéroporté (urédospores). Ce pathogène est unique parmi les rouilles car il possède de nombreux hôtes alternatifs, qui peuvent servir de sources d'inoculum. Les hôtes alternatifs sont d'autres plantes qui peuvent être infectées par le même pathogène, mais qui ne sont pas nécessaires pour compléter le cycle de vie du pathogène. Les hôtes alternatifs ne doivent pas être confondus avec l'hôte alternatif, qui est une plante autre que l'hôte principal, nécessaire à l'accomplissement du cycle de vie d'un pathogène. Dans les régions sans gel, comme l'Amérique du Sud, l'Amérique centrale, le bassin des Caraïbes, le sud du Texas et la Floride, la source d'inoculum pourrait se trouver à proximité sur des plants de soja spontanés, du kudzu ou un autre hôte alternatif. Dans les régions qui connaissent des gelées, comme le Midwest des États-Unis, l'inoculum doit être soufflé depuis des sources d'hivernage qui peuvent se trouver à des centaines de kilomètres.
La réintroduction d'agents pathogènes obligatoires dans une région éloignée se produit avec plusieurs autres maladies, comme la rouille de la tige du blé et les mildious, par exemple la moisissure bleue du tabac. Comme les spores de P. pachyrhizi sont sensibles aux rayons ultraviolets, le déplacement sur de longues distances de ces spores de rouille se produit probablement dans les systèmes de tempête où les nuages protègent les spores du soleil. Une fois que les spores viables se sont posées sur la surface des feuilles d'un hôte approprié, l'infection et le développement ultérieur de l'épidémie dépendent des conditions environnementales. En général, l'infection se produit lorsque les feuilles sont humides et que la température se situe entre 8°C et 28°C, avec un optimum entre 16°C et 28°C. À 25°C, une certaine infection se produit en seulement 6 heures de mouillage des feuilles, mais 12 heures sont optimales. Après l'infection, des lésions et des pustules avec des urédospores peuvent apparaître en 7 ou 8 jours, et le prochain cycle d'infection est prêt à commencer.
Ce cycle de vie court signifie que, dans des conditions favorables, les épidémies de rouille du soja peuvent rapidement passer de niveaux presque indétectables à des niveaux très élevés. Les épidémies de rouille du soja peuvent passer d'un niveau indétectable à la défoliation en un mois. Les épidémies peuvent sembler progresser encore plus vite que cela, car les premières infections se produisent dans la partie inférieure de la canopée et sont difficiles à trouver. Outre l'environnement, l'âge des plantes a une incidence sur les épidémies de rouille du soja. Habituellement, les lésions de rouille ne sont pas découvertes sur le soja avant la floraison, à moins que les niveaux d'inoculum ne soient élevés en début de saison. Cela peut être dû à une plus grande sensibilité des plantes à la rouille lorsque l'hôte entre dans les stades de reproduction, cela peut être dû au fait que dans les parties inférieures de la canopée les spores sont plus protégées des rayons UV, ou cela peut être dû au fait que les conditions dans la canopée deviennent plus humides à mesure que la canopée se referme. Dans tous les cas, des lésions peuvent se former à n'importe quel stade de croissance, mais les augmentations majeures de la maladie ne se produisent qu'après la floraison.
Deux modèles pour la rouille du soja sont décrits et calculés dans FieldClimate :
Modèle 1 : Les principaux facteurs pour le calcul de l'infection et de la gravité de la rouille du soja sont l'humidité des feuilles et la température (attention, l'infection doit être de 100% pour que les taux de gravité (de 1 à 3) commencent à être calculés). Les feuilles doivent être humides et la température doit être comprise entre 8 et 28°C - l'infection commence à être calculée (la somme de 7200 °minutes est de 100%). Si l'infection est de 100% et en fonction de la température et de la durée de mouillage des feuilles, les classes de gravité de 1 à 3 commencent à être calculées.
Modèle 2 : Ici, beaucoup plus de facteurs sont inclus dans le calcul de la rouille du soja : La température, l'humidité des feuilles, l'humidité relative, les précipitations et le rayonnement solaire (les hyphes fongiques peuvent pénétrer dans les stomates pendant la journée).
Lorsque l'infection de la rouille du soja atteint 100%, la sévérité commence à être calculée. Si l'infection et la gravité atteignent toutes deux 100%, l'apparition (l'établissement ?) d'un inoculum local est prévue.
La littérature :
- Ivancovich, A. y Botta, G. 2002. La roya de la sojo en la Argentinia. EEA Pergamino. RevistadeTEchnologia, Agropecuaria Vol. 7 (21), pp 16-17.
- Melching, J. S., Dowler, W. M., Koogle, D. L., et Royer, M. H. 1989. Effects of duration, frequency, and temperature of leaf wetness periods on soybean rust. Plant Dis. 73:117-122.
- X.B. Yang, E.M. Del Ponte, et A.P. Dias. Connaître le risque de rouille du soja par comparaison avec le Brésil. Département de phytopathologie, Université d'État de l'Iowa.
- Yang, X. B. 1995. Évaluation et gestion du risque de la rouille du soja. Compte rendu de l'atelier sur la rouille du soja, 9-11 août 1995. J. B.Sinclair et G. L. Hartman, eds. National Soybean Research Laboratory, Urbana, IL.
La pourriture sclérotique de la tige ou moisissure blanche du soja est une maladie causée par le champignon Sclerotinia sclerotiorum. La maladie peut entraîner des réductions importantes du rendement des semences, voire détruire complètement une culture lorsque le soja est planté dans un sol infesté et qu'il y a un couvert végétal dense avec des périodes prolongées de temps humide. Les pertes de rendement se produisent généralement lorsque l'incidence de la maladie est de 15 % ou plus. La plupart du temps, dans tout le Dakota du Nord, la pourriture de la tige causée par Sclerotinia ne constitue qu'un problème mineur à modéré pour les producteurs, et la maladie est rarement observée pendant les années de sécheresse. Le temps humide est un facteur important dans le développement de la maladie. Elle a été un problème persistant dans le soja irrigué. Outre la réduction du rendement des semences, la maladie entraîne également une diminution de la qualité des semences et une contamination des semences par les sclérotes noirs du champignon. La contamination des semences peut constituer un grave problème pour les semences exportées, car elle peut entraîner le rejet du lot de semences dans les ports d'entrée étrangers. En outre, les sclérotes retournés dans le sol peuvent affecter d'autres cultures dans la rotation. La pourriture sclérotique de la tige peut être gérée par les producteurs grâce à une bonne compréhension du pathogène et du cycle de la maladie.
Symptômes
Les symptômes ne sont généralement pas observés tant que le couvert végétal entre les rangs ne s'est pas refermé, créant un microclimat humide. Le flétrissement et le dessèchement des feuilles suivis de la mort des plantes sont généralement les premiers symptômes observés. Une inspection minutieuse sous le couvert végétal révèle une croissance mycélienne (fils de champignon) cotonneuse et blanche sur les tiges, les feuilles ou les gousses. Les lésions se développent sur les tiges principales et les branches latérales. Les lésions finissent par ceinturer les tiges et les parties de la plante situées au-dessus meurent. Les tiges sont blanchies et parfois déchiquetées par la pourriture avancée. De gros sclérotes noirs de formes et de tailles variées se forment à partir du mycélium blanc qui se développe sur les tissus végétaux. Les sclérotes se forment également dans la moelle de la tige et ont une forme cylindrique caractéristique. Les graines des gousses malades sont généralement ratatinées et peuvent être infectées par le champignon, ou remplacées par des sclérotes noirs. Les graines sont généralement contaminées par les sclérotes lorsque les plantes infectées sont récoltées.
Biologie des agents pathogènes
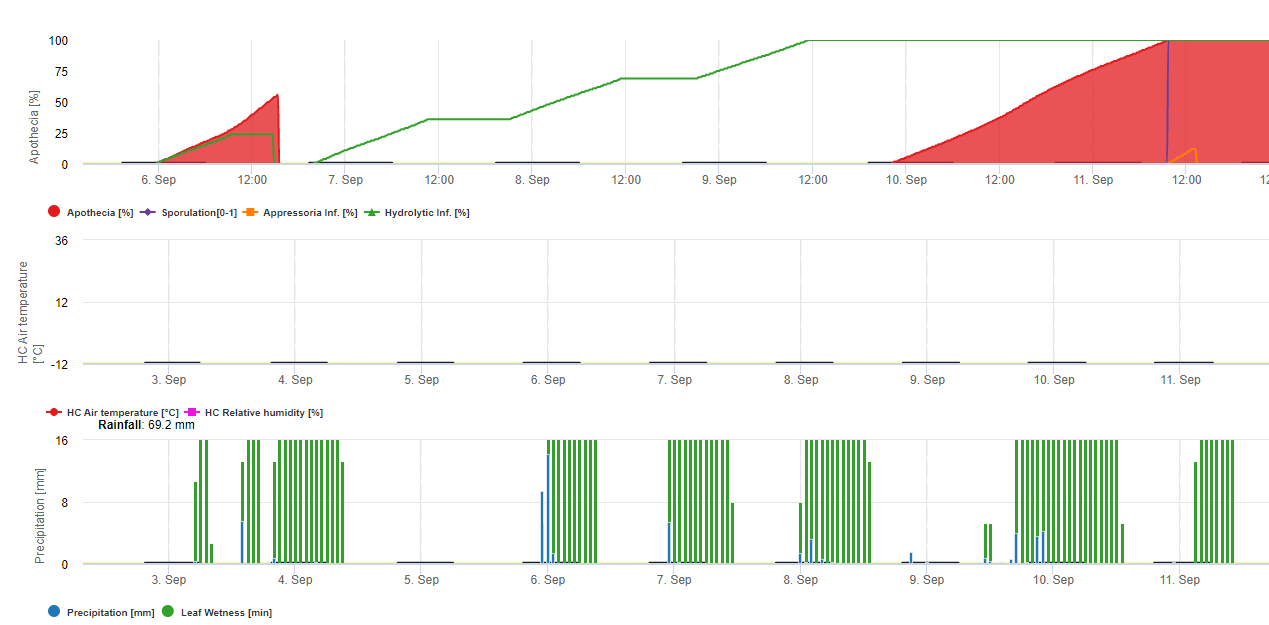
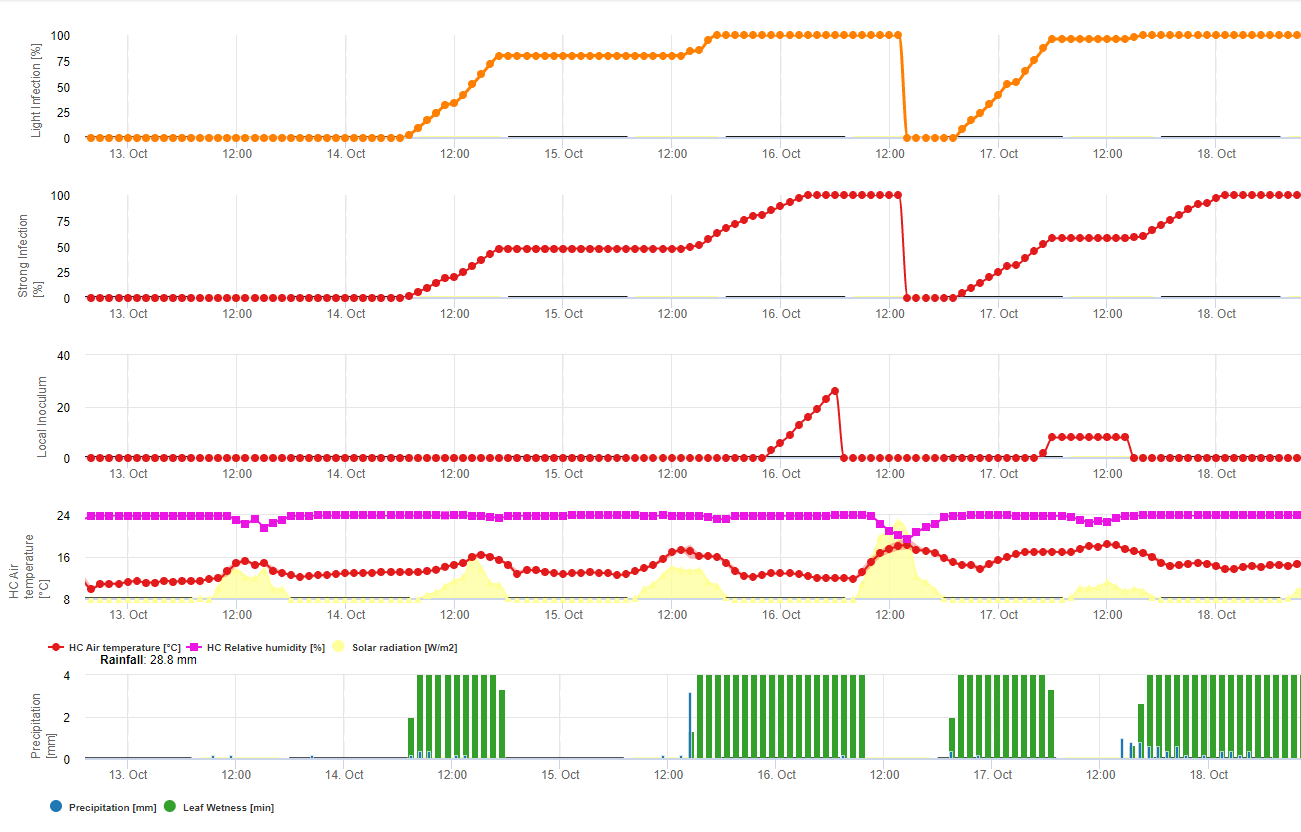
Ce champignon a une gamme d'hôtes étendue de plus de 370 espèces de plantes et provoque des maladies sur une grande variété de cultures telles que le tournesol, le haricot sec, le canola (colza), la pomme de terre, la luzerne, le sarrasin, le lupin, la moutarde, le topinambour, le carthame, la lentille, le lin, le pois de terre et de nombreux légumes. Dans le Dakota du Nord, il est rare que ce pathogène cause des dommages importants sur certaines de ces cultures, comme le lin et la pomme de terre. Il existe également de nombreux hôtes communs de mauvaises herbes à feuilles larges, comme le sureau des marais, le sarrasin, l'amarante, le chardon des champs et la moutarde sauvage. Le champignon qui cause la moisissure blanche sur le soja est le même que celui qui cause la moisissure blanche ou la maladie de Sclerotinia sur le tournesol, les haricots secs, le canola et d'autres cultures. Sclerotinia sclerotiorum passe l'hiver principalement sous forme de sclérotes dans le sol. Les sclérotes germent pour former de petits champignons bruns appelés apothécies (d'un huitième à un quart de pouce de diamètre). Ceux-ci produisent des spores appelées ascospores qui déclenchent la maladie sur le soja et d'autres cultures sensibles.
Cycle de la maladie
L'humidité et la floraison sont des facteurs critiques dans le développement des maladies. La maladie ne se déclare généralement pas avant la fermeture du couvert végétal, car un couvert dense favorise les températures fraîches et un microclimat humide autour des tiges et maintient une humidité élevée du sol après la pluie ou l'irrigation. L'initiation de la maladie est également étroitement associée à la floraison. Après sept à quatorze jours de forte humidité du sol, les sclérotes qui se trouvent dans les premiers centimètres du sol germent pour former des apothécies ressemblant à des champignons. Un seul sclérote peut produire plusieurs apothécies. Les apothécies vont éjecter de force leurs ascospores dans l'air où elles seront transportées par les courants d'air vers les plants de soja. La source la plus importante d'ascospores provient des apothécies produites dans le champ, mais les ascospores peuvent être transportées par le vent depuis des champs adjacents ou proches. Une apothécie peut produire d'énormes quantités d'ascospores sur plusieurs jours. Les ascospores survivent pendant de courtes périodes sur les tissus végétaux mais ne passent pas l'hiver. Les ascospores ont besoin d'un film d'eau et d'une base alimentaire telle que des tissus floraux morts ou en voie de sénescence pour germer et se développer avant d'infecter la plante. Les tissus floraux sont la base alimentaire la plus importante pour initier les infections. Les infections commencent souvent à l'aisselle des tiges où les tissus floraux sénescents sont tombés et se sont logés. Les infections peuvent également se produire à travers des blessures causées par la grêle ou d'autres dommages. Une pellicule d'eau à la surface de la plante favorise le développement des lésions et augmente l'étendue des dommages aux tissus. Le développement initial de la maladie nécessite généralement plus de 40 heures d'humidité continue à la surface de la plante, mais une fois que la maladie a commencé, des périodes d'humidité plus courtes permettent le développement des lésions. C'est la raison pour laquelle la maladie est associée à de longues périodes de temps nuageux, humide et pluvieux. Lorsque les surfaces des plantes sèchent, la progression de la maladie est ralentie. Les températures fraîches entre 59 et 75 degrés Fahrenheit favorisent le développement de la maladie. Plus le couvert végétal est dense, plus les conditions environnementales sont favorables à la maladie. Les sojas à semis solides et les cultures logées sont donc plus propices au développement de la maladie. Des sclérotes se formeront à mesure que le mycélium se développera dans et sur les tissus végétaux. Ces sclérotes ne germeront pas pour former d'autres apothécies au cours de la saison, mais retourneront plutôt dans le sol pendant la récolte et le travail du sol et passeront l'hiver pour devenir un inoculum (source de champignon infectieux) pour une future culture sensible. Les sclérotes sont des structures très résistantes et survivent pendant de longues périodes dans le sol.
Source : http://www.ndsu.edu/pubweb/~bernelso/soydiseases/sclerotinia.shtml
Dans FieldClimate, le modèle pour Sclerotinia est calculé en fonction des périodes de pluie, de l'humidité relative et de la température ainsi que de l'humidité des feuilles. Pendant les longues périodes humides, une infection par l'établissement d'un appressorium par le champignon pathogène est recommandée. Un autre mode d'infection est appelé "infection hydrolytique". Ce mode est basé sur la libération d'enzymes hydrolytiques qui dégradent séquentiellement la cuticule de la plante, les lamelles moyennes et les parois cellulaires primaires et secondaires, et rendent la plante entière. Modèle décrit en détail sur cette page.