

Pathogen
Pyricularia grisea, também conhecida como Magnaporthe oryzaeé um agente patogénico fúngico responsável pelo míldio do trigo, uma doença importante que afecta as culturas de trigo. O agente patogénico reproduz-se principalmente de forma assexuada através de conídios (esporos assexuados), que servem de inóculo primário para iniciar as infecções. Estes conídios são produzidos em conidióforos e são dispersos pelo vento, pela chuva e por meios mecânicos, como o equipamento agrícola. Ao pousar num hospedeiro de trigo suscetível, os conídios germinam e formam apressórios - estruturas especializadas que facilitam a penetração no tecido da planta. Uma vez lá dentro, o fungo coloniza as células do hospedeiro, levando ao desenvolvimento da doença. As condições ambientais desempenham um papel crucial no processo de infeção de P. grisea. O fungo desenvolve-se em climas quentes e húmidos, com temperaturas que variam entre 15°C e 30°C e períodos prolongados de humidade foliar ou humidade relativa elevada. A humidade contínua das folhas durante, pelo menos, 14 horas é necessária para iniciar a infeção, o que faz com que o excesso de nuvens, a humidade elevada, a irrigação excessiva e a irrigação inadequada favoreçam o desenvolvimento da doença. Além disso, factores de stress como a seca, a compactação do solo, as baixas alturas de corte e a fertilização excessiva com azoto podem exacerbar a gravidade da doença.
Sintomas
Os sintomas iniciais do míldio do trigo manifestam-se como lesões encharcadas de água, em forma de diamante, nas folhas, que depois se tornam cinzentas à medida que a doença progride. Estas lesões podem expandir-se e coalescer, conduzindo a uma extensa praga e necrose das folhas. As espigas infectadas exibem um branqueamento parcial ou total, muitas vezes a partir de um ponto de infeção cinzento-escuro na ráquis ou na base das espigas. Sob alta pressão de inóculo, podem ocorrer vários pontos de infeção numa única ráquis, resultando em perdas significativas de rendimento. Para além dos sintomas foliares, P. grisea pode infetar outras partes da planta de trigo acima do solo, incluindo os nós e o colo, provocando a podridão do colo ou a explosão dos nós. Esta situação pode provocar a rutura das partes afectadas, o que resulta em acamamento e maior redução do rendimento. A capacidade do agente patogénico de infetar várias partes da planta ao longo da estação de crescimento torna-o particularmente destrutivo, especialmente em condições ambientais favoráveis.
Modelo FieldClimate
Sensores necessários:
- Temperatura do ar
- Humidade relativa
- Molhagem de Folhas
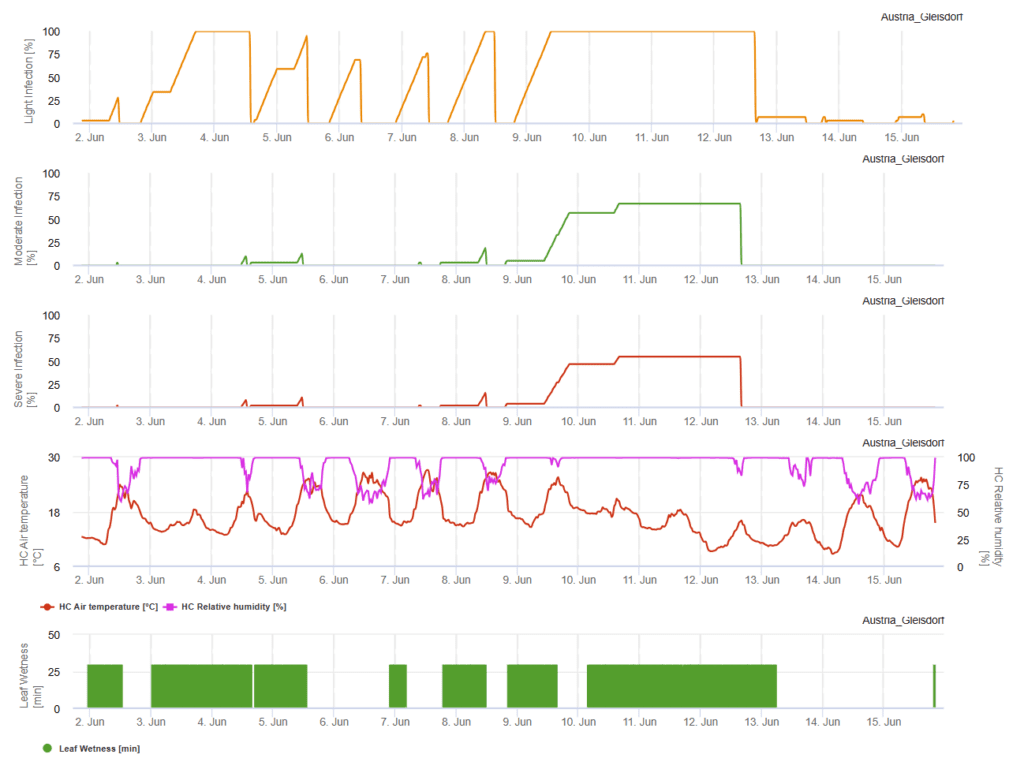
O céu nublado, a humidade e os chuviscos favorecem o desenvolvimento e a gravidade da doença. Em FieldClimate calculamos três classes de severidade e quando são atingidas 100% de infeção, as condições óptimas para o fungo patogénico são dadas no campo. Dependendo da variedade, o historial de campo e as medidas de proteção fitossanitária curativa devem ser tidos em conta quando uma infeção ligeira, moderada ou grave atinge 100%.
Literatura
- Cruz, C. D., & Valent, B. (2017). Doença da explosão do trigo: perigo em movimento. Patologia Vegetal Tropical, 42(3), 210-222.
- Islam, M. T., Croll, D., Gladieux, P., Soanes, D. M., Persoons, A., Bhattacharjee, P., ... & Talbot, N. J. (2016). O surgimento da explosão do trigo em Bangladesh foi causado por uma linhagem sul-americana de Magnaporthe oryzae. BMC Biology, 14(1), 84.
Pathogen
Pyrenophora teres (Drechslera teres) existe em duas formas - P. teres f. teres e P. teres f. maculata que induzem a mancha líquida em forma de rede e a mancha líquida em forma de mancha, respetivamente.
Os ascósporos libertados pela pseudoteca infectam os resíduos, dando início à infeção primária. Nalguns casos, os micélios e os conídios transmitidos pelas sementes também servem de inóculo primário. Os esporos germinam nas folhas e penetram na parede celular epidérmica externa para que o agente patogénico se possa desenvolver numa grande vesícula intracelular. Segue-se a fase assexuada, durante a qual são produzidos conídios que desencadeiam a infeção secundária, aumentando a gravidade da doença. No final da estação de crescimento, o agente patogénico produz pseudotecas como fonte de sobre-sementeira, que permanecem nos restos de cevada infectados.
Sintomas
Os sintomas manifestam-se nas folhas, caules e grãos, com a doença a progredir da base para o topo das plantas. As plantas mais velhas tendem a sofrer danos menos graves, uma vez que possuem uma cutícula mais espessa que impede a penetração do agente patogénico e uma maior capacidade de produzir compostos antifúngicos. Os agentes patogénicos produzem toxinas que são responsáveis pelos sintomas, contribuindo para a necrose e a clorose ou perturbando o equilíbrio hídrico a nível celular.
No caso da doença da mancha em rede, os sintomas aparecem como lesões estreitas, castanho-escuras e longitudinais. Nas variedades altamente resistentes, desenvolver-se-ão apenas algumas pequenas lesões pontilhadas, sem formação de um padrão distinto em forma de rede. Em contrapartida, as variedades mais susceptíveis podem apresentar áreas cloróticas ou encharcadas de água à volta das lesões necróticas.
No caso da mancha, os sintomas consistem em lesões castanho-escuras, circulares ou elípticas, frequentemente rodeadas por uma auréola clorótica ou necrótica de largura variável. Nas variedades menos susceptíveis, as lesões são geralmente mais pequenas e podem não ter uma auréola clorótica circundante.
As infecções graves podem resultar na morte completa das folhas, dando-lhes um aspeto seco. As folhas mais velhas normalmente apresentam primeiro a murchidão, seguidas pelas folhas mais jovens.
Modelo FieldClimate
Drechslera teres Modelo
Sensores necessários:
- Temperatura do ar
- Humidade relativa
- Molhagem de Folhas
- Precipitação
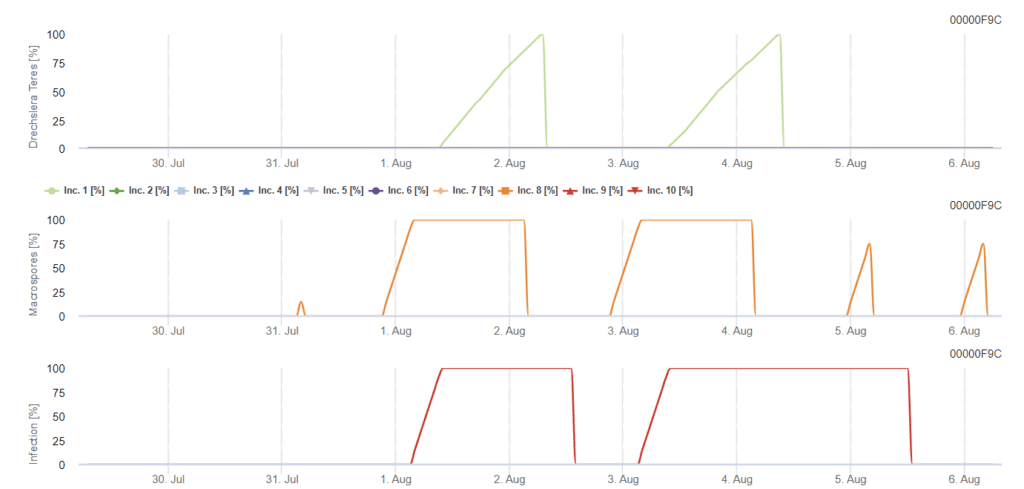
O inóculo primário desenvolve-se a temperaturas óptimas entre 15°C e 25°C, sendo a óptima de 20°C. Condições superiores a 6 horas de escuridão conduzem a um crescimento ótimo dos conídios, como se pode ver num gráfico de esporulação de conídios. Se o gráfico de esporulação atingir 100%, as condições óptimas são estabelecidas no campo.
A dispersão posterior ocorre através do vento ou da chuva. As infecções dos tecidos vegetais necessitam de condições de humidade, tais como humidade foliar ou humidade relativa elevada, durante cerca de 10 a 30 horas, com temperaturas óptimas de 15°C e 25°C, como se pode ver num gráfico de infeção. Quando atinge 100%, determina-se que foram medidas as condições óptimas para a infeção no campo. A luz solar também pode aumentar o crescimento de conidióforos que produzem conídios, seguindo condições alteradas de períodos húmidos e secos com alta radiação solar e baixa humidade relativa, e o vento favorece o desenvolvimento da doença.
Literatura
- Backes, A., Guerriero, G., Ait Barka, E., & Jacquard, C. (2021). Pyrenophora teres: taxonomia, morfologia, interação com a cevada e modo de controlo. Fronteiras na ciência das plantas, 12, 614951.
- Liu, Z., Ellwood, S. R., Oliver, R. P., & Friesen, T. L. (2011). Pyrenophora teres: perfil de um agente patogénico da cevada cada vez mais prejudicial. Patologia molecular das plantas, 12(1), 1-19.
- Obst, A., & Paul, VH (1993). Doenças e pragas dos cereais, Verlag Th. Mann, Gelsenkirchen-Buer.
Pathogen
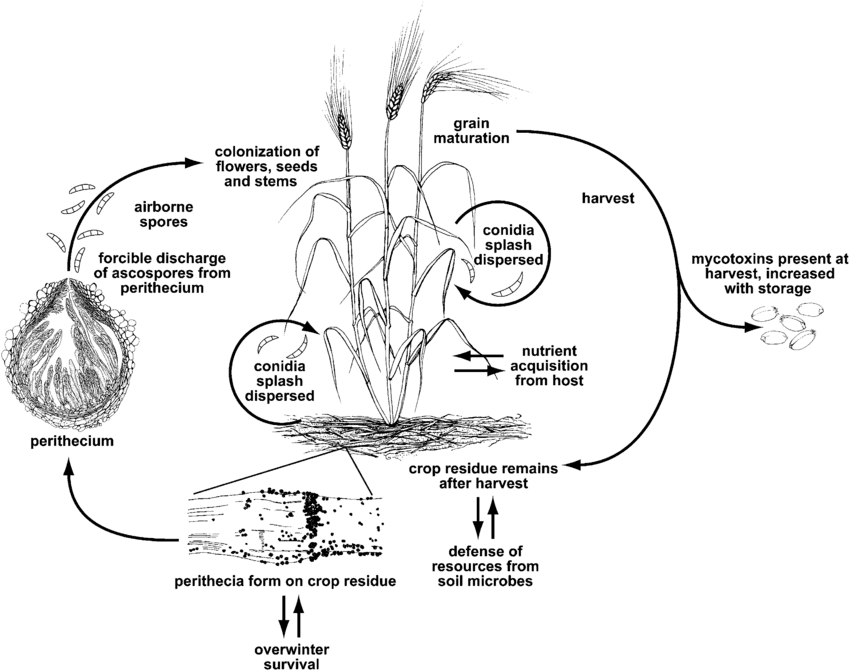
O míldio ou sarna de Fusarium é causado pelo género Fusarium. A espécie mais comum é Fusarium graminearum mas as espécies mais prevalecentes podem diferir consoante as regiões.
F. graminearum pode infetar plantas de trigo em qualquer fase de crescimento. Sobrevive ao inverno em resíduos de culturas infectadas e, na primavera, a infeção primária ocorre quando os ascósporos dos peritécios e conídios se dispersam nas glumas ou anteras do trigo. As florzinhas infectadas produzem grãos murchos ou enrugados, frequentemente designados por grãos "pedra tumular". Mesmo quando os grãos não parecem afectados, podem ainda estar contaminados com micotoxinas. A infeção secundária ocorre através de conídios, mas as epidemias de trigo dependem principalmente da quantidade de inóculo primário e não de fontes secundárias.
O míldio de Fusarium também leva à acumulação de micotoxinas nocivas nas culturas, constituindo um risco potencial para os animais. Estas micotoxinas desactivam o mecanismo de defesa das plantas ou protegem o fungo contra outros organismos. A principal toxina é o desoxinivalenol (DON), que inibe a síntese proteica, perturbando o funcionamento normal das células. Os seres humanos que consumiram trigo com DON apresentam sintomas como náuseas, febre ou vómitos.
Sintomas

No trigo e no trigo duro, qualquer parte ou a totalidade da cabeça pode aparecer branqueada. As cabeças parcialmente brancas e verdes são o diagnóstico da doença no trigo. O fungo pode também infetar o caule, causando uma descoloração castanha a púrpura. Massas de esporos cor-de-rosa a laranja-salmão são frequentemente observadas nas espiguetas e glumas infectadas em condições de humidade. Muitos grãos estão enrugados e leves e são por vezes chamados "pedras tumulares" devido ao seu aspeto calcário e sem vida. Em condições favoráveis, a infeção pode espalhar-se para as espiguetas adjacentes e infetar toda a espiga, incluindo a ráquis e o pedúnculo. Os grãos podem ter um aspeto baço e uma descoloração rosada em tamanho normal quando a infeção ocorre numa fase tardia do desenvolvimento.
Na cevada, as espiguetas infectadas ficam branqueadas e apresentam um aspeto encharcado. Os grãos também podem apresentar descoloração e, ocasionalmente, podem ser observadas massas de esporos de cor laranja-salmão na espigueta e nas glumas em condições de humidade.
Modelos FieldClimate
As estimativas dos níveis de FHB num campo baseiam-se na contagem de cabeças ou grãos atacados. A escala visual colorida para estimar a severidade da FHB no trigo está disponível no Serviço de Extensão da NDSU (publicação PP-1095): https://library.ndsu.edu/ir/bitstream/handle/10365/9187/PP1095_1998.pdf?sequence=1&isAllowed=y.
Modelo de Infeção por Fusarium Head Blight (Risco de Fusarium Head Blight)
Sensores necessários:
- Temperatura do ar
- Humidade relativa
- Humidade das folhas
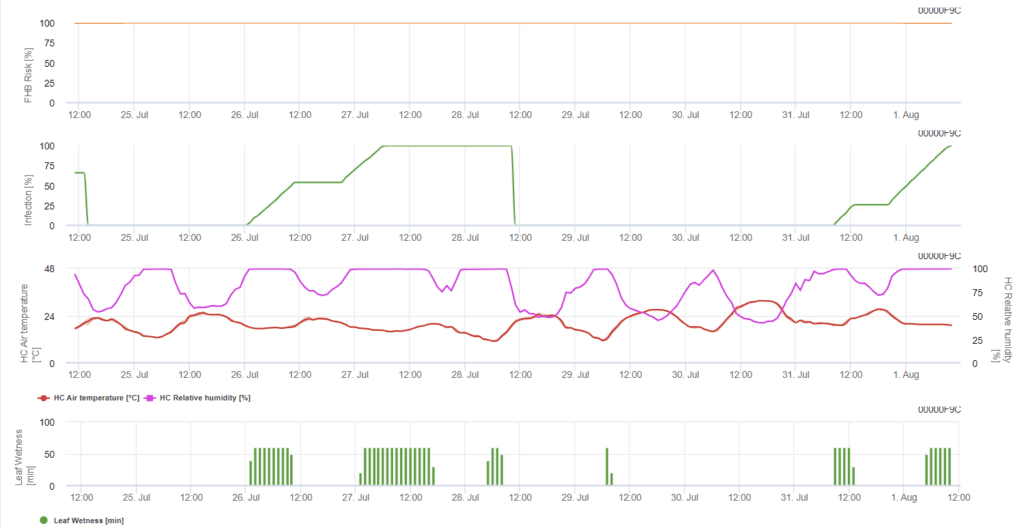
O modelo determina os períodos de risco para uma infeção. Sempre que se atinge a infeção 100%, o risco é muito elevado e as condições para o fungo são favoráveis à infeção. O conhecimento do agricultor sobre o estádio de desenvolvimento das diferentes variedades de trigo dá-lhe a possibilidade de decidir a aplicação de uma pulverização curativa imediatamente após a infeção.
Os agentes fúngicos do Fusarium Head Blight no trigo são favorecidos por temperaturas quentes de 20°C a 30°C e longos períodos de humidade. Vários períodos de humidade nas folhas, que duram um dia, conduzem a sintomas visíveis precocemente. Os sintomas também podem ser vistos após um longo período de latência se a infeção for seguida por um período de humidade da folha de 18 horas ou mesmo mais curto e após um inóculo artificial a temperaturas de 15°C.
Resumindo todas as diferentes combinações de temperatura e humidade que encontrámos na literatura, decidimos assinalar Infecções por Fusarium Head blight se a temperatura e o período de humidade foliar ou períodos com mais de 85% de humidade relativa excederem os valores visualizados no gráfico seguinte. O cálculo do valor de progresso segue a relação entre a duração das condições de humidade e a temperatura.
Modelo de alerta para micotoxinas de Fusarium (Fusarium Head Blight)
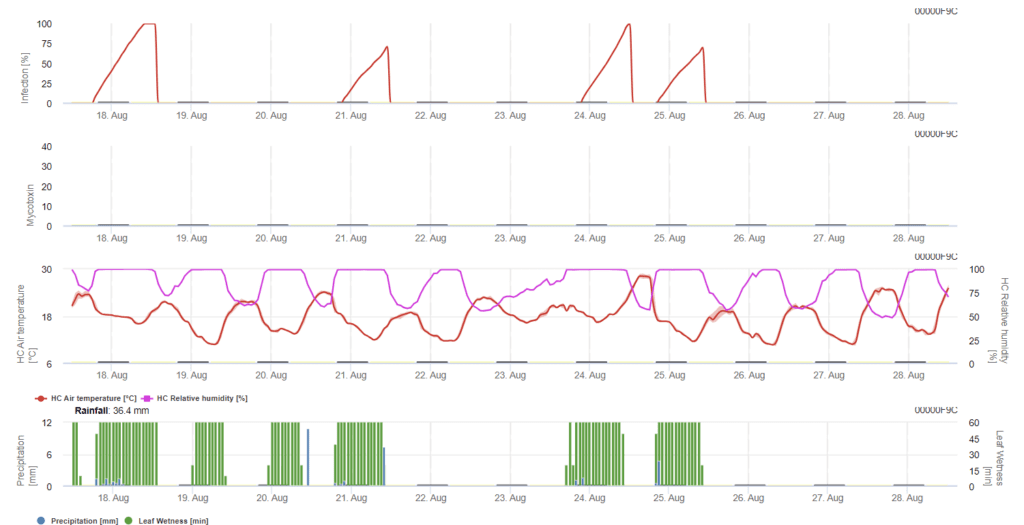
Sensores necessários:
- Temperatura do ar
- Humidade relativa
- Humidade das folhas
- Precipitação
neste modelo a infecção da FHB é calculada por precipitação (2mm necessários), humidade relativa (acima de 85%) ou humidade da folha, temperatura durante o processo. Se a infecção atingir 100%, as condições óptimas para o agente patogénico fúngico foram atingidas. Mais adiante, o modelo calcula o risco de micotoxinas FHB.
Os ensaios de infeção com períodos prolongados de humidade foliar do Fusarium head blight conduzem a elevados teores de micotoxinas. A partir desta informação, presume-se que um período de humidade das folhas de 48 horas ou mais durante os estádios 61 e 69 representa um risco elevado de micotoxinas.
A experiência adquirida durante a análise do DON no trigo cultivado para fins comerciais mostrou que os períodos de humidade das folhas suficientemente longos para a infeção, após uma infeção inicial nos estádios 61 a 69, podem aumentar os valores de DON. No caso de períodos de humidade foliar mais longos, as micotoxinas podem aumentar até ao estádio 85.
- Estágio 61 de BBCH: Início da floração; primeiras anteras visíveis
- Estádio 69 de BBCH: Fim da floração; todas as espiguetas completaram a floração, mas podem permanecer algumas anteras desidratadas
- BBCH estádio 85: A meio da maturação; massa macia com conteúdo de grãos macios mas secos, impressão da unha não mantida
O modelo acumula o valor do risco proporcional ao progresso da infeção para cada período de infeção bem sucedida durante o período selecionado para o cálculo. Seis infecções completas conduziriam a um risco de 100%. Normalmente, o período de humidade das folhas que conduz a uma infeção por fusarium é mais longo do que o mínimo necessário. Por conseguinte, a maioria das infecções por fusarium conduzirá a um aumento do risco superior a 17%.
O valor do risco de micotoxinas depende do historial do campo. O trigo cultivado após o plantio direto só pode ter um pequeno risco se não for pulverizado na situação ideal. No trigo não pulverizado, é de esperar um aumento dos valores de DON após 35% do risco. O trigo após o trigo não lavrado que se segue a qualquer outra cultura pode acarretar um risco mais elevado de 50%. Se tivermos trigo após trigo com lavoura, o risco pode aumentar até 70%. O trigo de primeiro ano deve ser testado para DON se o risco atingir 100%.
Literatura
- https://www.fao.org/4/y4011e/y4011e0j.htm
- https://scabusa.org/pdfs/NDSU_PP-804_FHB-Small-Grains.pdf
- Lancashire, P. D., Bleiholder, H., Boom, T. V. D., Langelüddeke, P., Stauss, R., Weber, E., & Witzenberger, A. (1991). Um código decimal uniforme para estádios de crescimento de culturas e infestantes. Anais de Biologia Aplicada, 119(3), 561-601.
- Trail, F. (2009). Por ondas de grãos atacados: Fusarium graminearum na era pós-genómica. Fisiologia vegetal, 149(1), 103-110.
- Schumann, G. L. (2010). O instrutor fitossanitário| Volume: 10| Ano: 2010| Tipo de artigo: Planos de aula. Fitossanidade, 10.
Pathogen
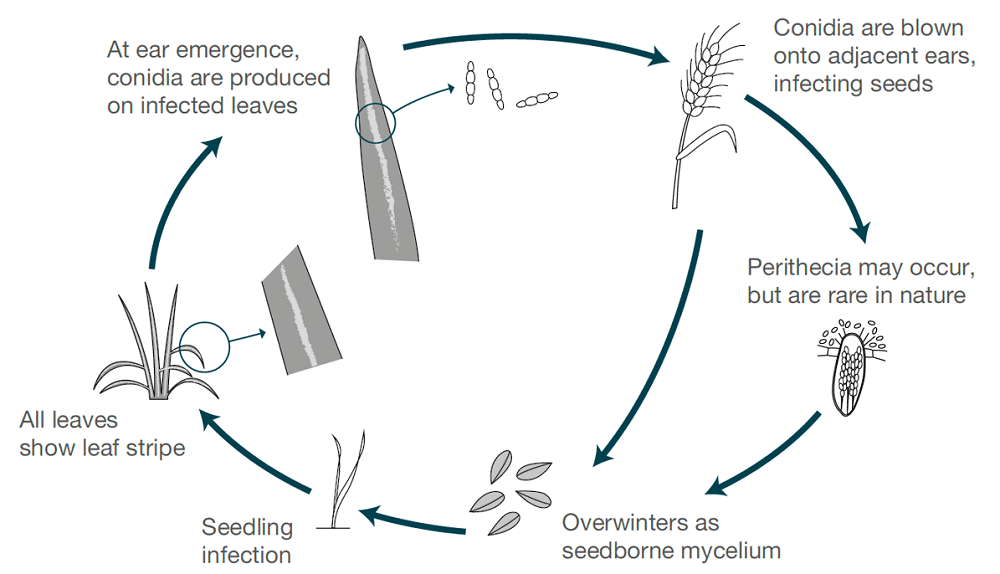
A risca de cevada é causada por Pyrenophora graminea (Drechslera graminea)O fungo da família das sementes, um agente patogénico transmitido por sementes que sobrevive na epiderme de uma semente infetada. O fungo infecta as plântulas em condições frescas e húmidas. O fungo cresce sistemicamente na planta, infecta toda a planta, produz toxinas, mata as células e descolora o tecido foliar entre as nervuras, causando assim lesões estriadas. A estria é mais frequente na cevada de inverno do que na cevada de primavera.
Quando as condições são húmidas ou molhadas, os esporos são produzidos na superfície das folhas quando os espinhos das plantas saudáveis florescem. Os esporos são dispersos pelo vento para estas espigas em desenvolvimento, germinam e causam infecções. Os grãos são mais susceptíveis durante o desenvolvimento inicial. Apenas um ciclo de infeção e produção de esporos ocorre em cada estação.
Sintomas

Os sintomas são mais proeminentes na fase de emergência da orelha.
Na parte basal da folha e da bainha aparecem inicialmente riscas verde-pálidas a amarelas. Estas riscas tornam-se gradualmente castanhas a castanho-escuras, seguidas de secagem e divisão da lâmina foliar. Estas estendem-se a todo o comprimento da folha, tornam-se necróticas e coalescem, levando eventualmente à morte da planta.
As plantas podem parecer raquíticas, produzir poucos perfilhos e, em casos graves, não produzir cabeça ou sementes. O comprimento das espigas pode também ser reduzido devido ao fraco desenvolvimento dos grãos castanhos.
Modelo FieldClimate
Pyrenophora graminea Modelo
Sensores necessários:
- Temperatura do ar
- Humidade relativa
- Humidade das folhas
- Precipitação
O modelo apresenta duas fases. A primeira fase corresponde a períodos de risco durante os períodos de emergência da planta, fases iniciais. A segunda fase é durante o período de floração e desenvolvimento da cabeça da cevada. Quando a infeção atinge 100%, estão criadas no campo as condições ideais para que o fungo infecte os tecidos da planta e cresça sistematicamente no seu interior.
Literatura
- Richardson, M. J. (1996). Micologia de sementes. Investigação micológica, 100(4), 385-392. https://ahdb.org.uk/knowledge-library/barley-leaf-stripe-life-cycle-and-disease-symptoms https://plantwiseplusknowledgebank.org/doi/full/10.1079/pwkb.species.46115
Pathogen
Existem duas doenças principais da septoriose no trigo - a mancha de Septoria tritici causada por Septoria tritici e a mancha de Septoria nodorum causada por Septoria nodorum.
Septoria tritici sobrevive em resíduos de culturas infectadas durante o verão e começa a infetar as plantas de trigo no outono. O fungo desenvolve-se em condições frescas e húmidas. Tem duas fases - uma fase latente e uma fase necrótica. Durante a fase latente, o fungo inocula novos tecidos vegetais e, entrando através dos estomas, segue-se a colonização, crescendo intercelularmente nos tecidos vegetais. S. tritici não necessita de quaisquer estruturas físicas de alimentação, mas alimenta-se extraindo nutrientes das células hospedeiras que perderam a integridade estrutural. As plantas podem apresentar uma resposta hipersensível e podem desenvolver-se lesões necróticas nos tecidos afectados como resposta à infeção.
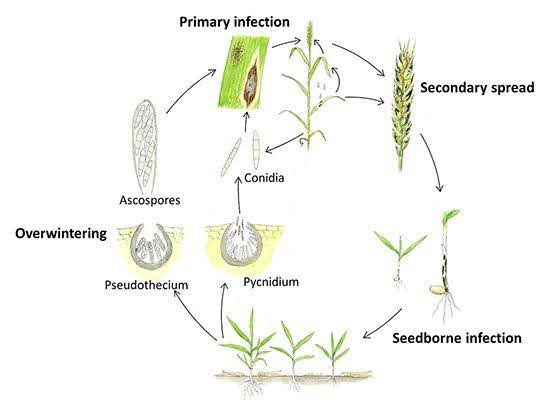
Septoria nodorum sobrevive durante o verão sob a forma de micélios dormentes, picnídios e pseudotecas em resíduos de culturas. A infeção começa normalmente no outono, quando os picnidiósporos dos picnídios e os ascósporos das pseudotecas são dispersos pelo vento e pela chuva, dando início à infeção primária. Estes esporos penetram na cutícula da folha e a formação de picnídios segue-se à infeção. A infeção secundária ocorre quando os picnidiósporos são produzidos e dispersos das folhas inferiores para as folhas superiores e glumas. O agente patogénico também produz toxinas que contribuem para o desenvolvimento da doença.
Sintomas
Mancha de Septoria tritici
No outono, Septoria tritici Os sintomas aparecem inicialmente como pequenas manchas amarelas nas folhas, contendo minúsculas manchas pretas, que são corpos de frutificação do fungo. As lesões têm uma forma irregular, variando de elípticas a longas e estreitas, aumentam de tamanho e tornam-se castanhas a castanhas avermelhadas à medida que amadurecem. A doença começa normalmente nas folhas inferiores e move-se gradualmente para cima, acabando por afetar a folha bandeira. Em condições de humidade, o fungo pode propagar-se às cabeças do trigo, causando lesões castanhas nas glumas e nos colmos, conhecidas por mancha das glumas. A mancha de Septoria tritici pode ser confundida com outras doenças do trigo. No entanto, a presença de corpos fúngicos negros é uma caraterística fundamental da S. tritici. Embora a Septoria nodorum também possa produzir corpos de frutificação castanhos em condições de humidade, estes diferem em cor e tamanho, com Septoria tritici os corpos de frutificação são maiores.
Mancha de Septoria nodorum
Septoria nodorum Os sintomas aparecem principalmente nas folhas superiores. Inicialmente, aparecem nas folhas manchas castanhas escuras com uma auréola amarela. A queimadura das pontas das folhas é outra indicação inicial. As lesões expandem-se e tornam-se necróticas com um centro castanho escuro à medida que amadurecem. No interior das lesões são visíveis pequenas estruturas castanhas escuras, que são corpos de frutificação. Os conídios libertados dos corpos de frutificação têm um aspeto branco a rosa. As folhas gravemente infectadas podem ficar completamente cobertas por lesões, resultando eventualmente na morte do tecido foliar. O fungo pode também afetar as glumas e os colmos: aparecem lesões castanhas semelhantes e a doença progride para baixo.
Modelo FieldClimate
As infecções por septórios são possíveis a baixas temperaturas, enquanto que temperaturas inferiores a 7°C podem não conduzir a uma infeção dentro de 2 dias. A temperatura óptima da doença é atingida na zona de 16 a 21°C. As infecções são possíveis durante um período de humidade relativa elevada ou de humidade foliar de 14 horas ou mais.
Septoria tritici Modelo
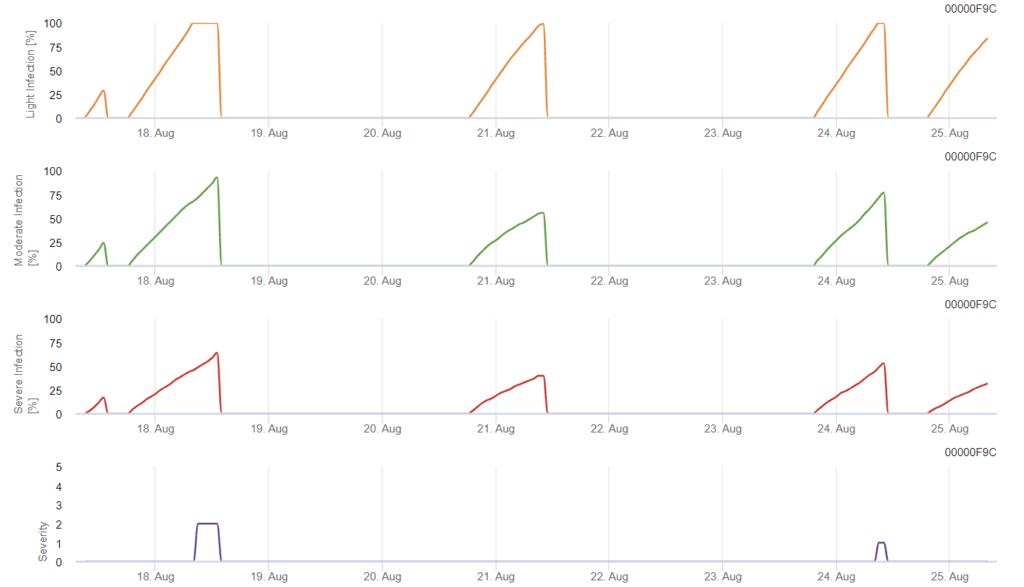
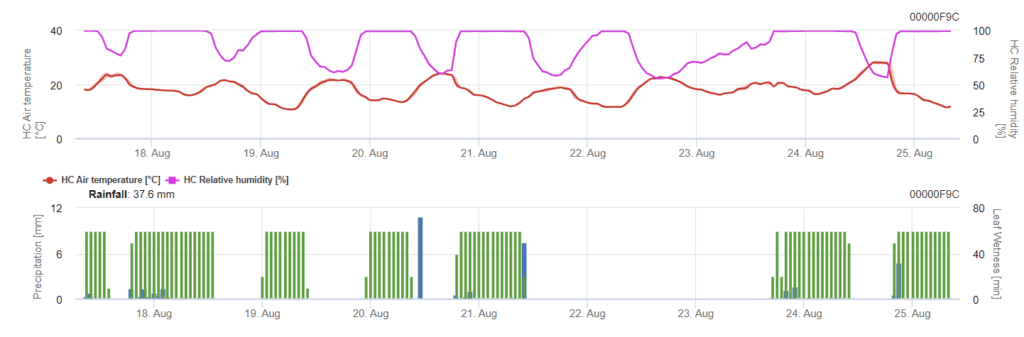
A infeção começa depois de uma chuva de 0,5 mm. As temperaturas ideais para as infecções por Septoria são cerca de 16°C a 21°C e humidade foliar ou humidade relativa elevada durante 14 horas. A Septoria tritici é calculada em três classes de severidade. Além disso, determinamos níveis de severidade que apoiam a avaliação da pressão de infeção entre os estádios BBCH 10 a 32 do trigo.
- BBCH fase 10: Desenvolvimento da folha; primeira folha através do coleóptilo
- BBCH estádio 32: Alongamento do caule; nó 2 pelo menos 2 cm acima do nó 1
- Estádio 51 de BBCH: Emergência da inflorescência e espigamento; início do espigamento com a ponta da inflorescência emergida da bainha e a primeira espigueta apenas visível.
Decidimos não utilizar um modelo para a formação de picnídios. A condição necessária para a formação de picnídios é assumida como sendo um período com humidade relativa superior a 85%. O tempo de vida dos picnídios é de 24 horas. Em todos os climas em que o fungo tem a possibilidade de infetar, encontraremos duas horas que preenchem esta condição quase todos os dias por volta do nascer do sol.
Avaliação da gravidade da infecção
Para avaliar o Septoria tritici pressão de infeção entre os estádios 10 e 32 e entre 32 e 51, temos de avaliar a gravidade das infecções com base nas condições climáticas. A gravidade da infeção é avaliada numa escala de 1 a 5:
- 1: infeção fraca com chuva inferior a 5 mm
- 2: infeção fraca com chuva superior a 5 mm
- 3: infeção moderada com chuva inferior a 5 mm
- 4: infeção moderada com chuva superior a 5 mm/ infeção grave com chuva inferior a 5 mm
- 5: infeção grave com chuva superior a 5 mm
Avaliação da pressão da doença
Existem três factores para a pressão da doença no campo - o clima, a história do campo e a suscetibilidade da variedade cultivada. Se pudermos acumular os valores de severidade da doença desde o estádio 10 até ao estádio 32 até ao valor de 4, podemos esperar uma fraca pressão da doença pelo clima. Se este valor atingir 6, podemos esperar uma pressão moderada da doença, e se atingir 10, uma pressão mais elevada da doença por parte do clima.
Conhecer a suscetibilidade da variedade e o historial do campo levar-nos-á a pulverizar ou não numa situação de pressão fraca ou moderada da doença. Um valor acumulado de 10 pode, de qualquer forma, levar a uma pulverização no estádio 32.
A decisão de pulverizar numa fase posterior depende do clima da primavera. Se pudermos acumular os valores de severidade do estádio 10 até um valor de 6, podemos esperar uma fraca pressão da doença. Se este valor atingir 10, podemos esperar uma pressão moderada da doença e se este valor atingir 15, podemos esperar uma pressão elevada da doença devido à situação climática.
Em FieldClimate, o Septoria tritici juntamente com as três diferentes severidades de infeção. Devido à precipitação e aos longos períodos de humidade das folhas, estão reunidas as condições para uma infeção grave por S. tritici. Os níveis de severidade atingem o valor mais elevado de 5, o que significa que existe um risco elevado de infeção.
Septoria nodorum Modelo
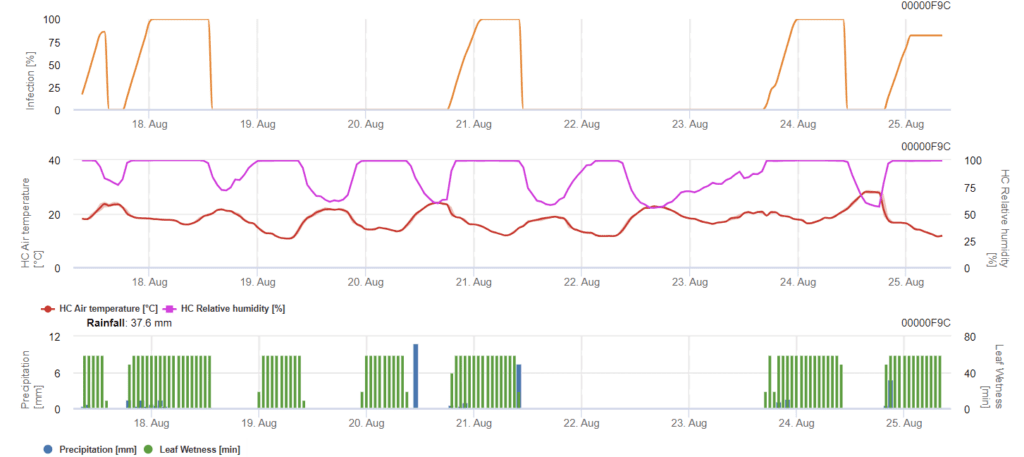
Septoria nodorumA biologia da infeção de uma pessoa difere em certa medida da Septoria tritici mas a diferença não é suficientemente grande para um modelo separado. Por conseguinte, sugerimos a utilização do modelo tritici para todo o complexo da doença de Septoria. Em zonas com elevada pressão de S. nodorumSe o vírus for classificado como fraco, com um valor de gravidade de 2, as infecções devem ser tratadas com mais seriedade do que noutras áreas.
O Septoria nodorum calcula o risco da doença. As medidas de proteção das plantas devem ser consideradas se o risco atingir 80%. Quando o risco é de 100% e a infeção já está determinada, devem ser tomadas medidas fitossanitárias sistémicas (aplicações curativas).
Literatura
- Brennan, C. J., Benbow, H. R., Mullins, E., & Doohan, F. M. (2019). Uma revisão das incógnitas conhecidas nos estágios iniciais da doença da mancha de septoria tritici do trigo. Fitopatologia, 68(8), 1427-1438.
- De Wolf, E. D. (2008). Septoria tritici blotch. Mehra, L. K., Adhikari, U., Ojiambo, P. S., & Cowger, C. (2019). Mancha de Septoria nodorum do trigo. O instrutor de fitossanidade.
- Solomon, P. S., Lowe, R. G., TAN, K. C., Waters, O. D., & Oliver, R. P. (2006). Stagonospora nodorum: causa da mancha de stagonospora nodorum do trigo. Patologia molecular das plantas, 7(3), 147-156.
Pathogen
A mancha da folha é causada por Rynchosporium secalis.
A infeção primária ocorre através de ascósporos ou conídios em restos de culturas. Estes esporos são dispersos pelo vento e pela chuva, germinam e, à medida que as células da epiderme e do mesófilo colapsam após uma infeção, os sintomas tornam-se visíveis. A infeção secundária é provocada por conídios provenientes de folhas infectadas. Em condições de humidade, os conídios germinam na superfície da folha e produzem hifas que penetram na cutícula diretamente acima das células epidérmicas. O crescimento subsequente do fungo está confinado à região subcuticular da epiderme.
Sintomas
R. secalis pode infetar qualquer parte das folhas. As manchas de forma irregular são a caraterística principal e a infeção começa da parte inferior para a parte superior das folhas. As manchas aparecem inicialmente como áreas encharcadas de água. À medida que a doença progride, tornam-se acinzentadas no centro devido à formação de esporos com uma margem castanha. Pode seguir-se necrose e clorose. As manchas coalescem, destruindo as folhas inteiras.
Modelo FieldClimate
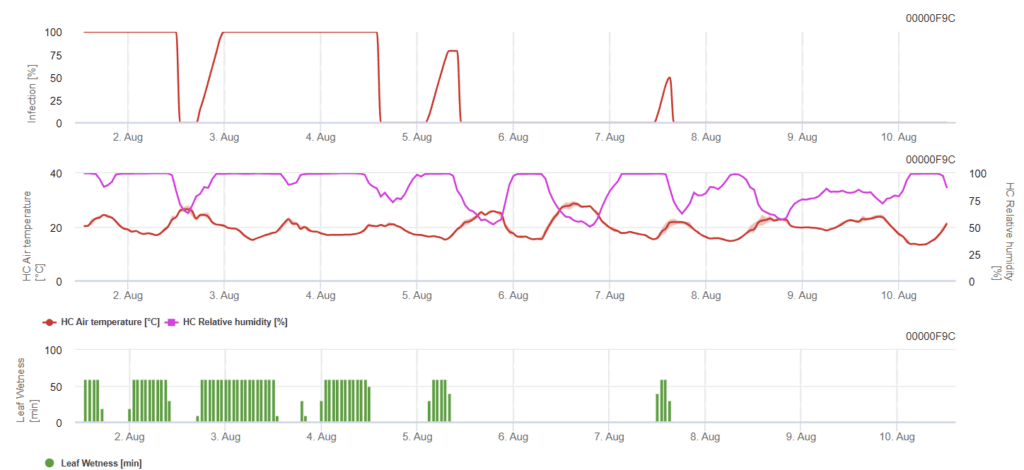
Modelo de Rynchosporium da mancha da folha
Sensores necessários:
- Temperatura do ar
- Humidade relativa
- Humidade das folhas
Para que ocorra uma infeção, é necessário um mínimo de 7 a 15 horas de humidade do chumbo, dependendo da temperatura (óptima a 20'C). Quando o gráfico atinge 100%, significa que as condições óptimas foram determinadas no campo.
Literatura
- Brooks, F. T. (1928). Observations on Rhynchosporium secalis (Oud.) Davis, leaf blotch of barley and rye. Novo Fitólogo, 27(4), 215-219.
- Fowler, A. M., & Owen, H. (1971). Estudos sobre a mancha foliar da cevada (Rhynchosporium secalis). Transactions of the British Mycological Society, 56(1), 137-152.
- Zhan, J., Fitt, B. D., Pinnschmidt, H. O., Oxley, S. J. P., & Newton, A. C. (2008). Resistência, epidemiologia e gestão sustentável das populações de Rhynchosporium secalis na cevada. Patologia vegetal, 57(1), 1-14.
Pathogen
Ramularia collo-cygniO fungo Ramularia, agente causal da mancha foliar da Ramularia na cevada, propaga-se principalmente por reprodução assexuada através de conídios, que requerem humidade para a germinação e desenvolvimento inicial. Após a germinação na superfície da folha, o fungo entra na folha através dos estomas e coloniza o tecido intercelularmente. As plantas de cevada infectadas podem não apresentar sintomas inicialmente, mas os danos aparecem normalmente após a floração.
O ciclo de vida de R. collo-cygni não é totalmente compreendido, mas envolve a transmissão vertical através de sementes infectadas, permitindo que o agente patogénico sobreviva entre estações de crescimento. Os conídios transportados pelo ar também contribuem para a propagação secundária, iniciando novas infecções em condições favoráveis. Embora as sementes sejam o principal mecanismo de invernada, o papel dos hospedeiros alternativos e de uma estrutura fúngica secundária denominada Asteromela permanece incerto e está ainda a ser estudado.
Sintomas

Os sintomas aparecem em ambos os lados das folhas após a floração, em geral, mas são mais comuns nas folhas superiores. Os sintomas iniciais aparecem como manchas amarelas a castanhas nas nervuras das folhas, rodeadas por uma auréola clorótica. Quando as manchas surgem, a folha torna-se clorótica e necrótica, começando normalmente na ponta e nas margens da folha. As manchas vizinhas podem encontrar-se, criando áreas escuras maiores. Podem também ser observadas pequenas manchas punctiformes. Embora os sintomas da mancha foliar de Ramularia possam ser confundidos com outras doenças, a caraterística principal é que as manchas foliares são delimitadas às nervuras da folha.
Existem os chamados 5R para distinguir as manchas foliares de Ramularia:
- Anelada com uma margem amarela de clorose
- Forma retangular
- Limitada pelas nervuras da folha
- Coloração castanho-avermelhada
- Mesmo através da folha
Modelo FieldClimate
Modelo Ramularia
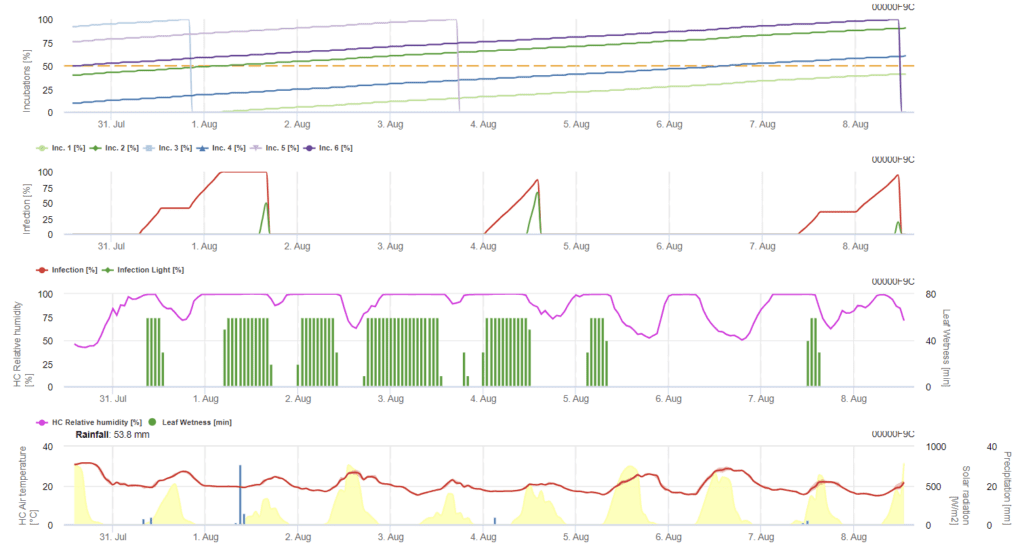
A infeção por Ramularia na cevada é principalmente causada por condições de humidade. Assim, iniciamos o cálculo com um evento de chuva, seguido de humidade foliar ou humidade relativa superior a 85%.
Existem duas formas de descrever uma infeção:
- Infeção: uma vez que as condições de humidade apenas promovem um evento de infeção
- Infeção: uma vez quando a radiação global é necessária para terminar um evento de infeção
Posteriormente, sempre que se atinge 100% de infeção, o modelo começa a determinar o tempo de incubação. Quando a incubação atinge 50%, recomenda-se o controlo do campo para novos inóculos e quando atinge 100%, prevê-se a ocorrência de lesões necróticas nas folhas.
Literatura
- Havis, N. D., Brown, J. K., Clemente, G., Frei, P., Jedryczka, M., Kaczmarek, J., ... & Hess, M. (2015). Ramularia collo-cygni-um patógeno emergente de culturas de cevada. Fitopatologia, 105(7), 895-904.
- Huss, H. (2002, abril). A biologia de Ramularia collo-cygni. In Proc. Second Int. Workshop Barley Leaf Blights. Aleppo, Síria (pp. 321-328).
- Walters, D. R., Havis, N. D., & Oxley, S. J. (2008). Ramularia collo-cygni: a biologia de um agente patogénico emergente da cevada. Cartas de Microbiologia FEMS, 279(1), 1-7. https://grdc.com.au/__data/assets/pdf_file/0025/443509/GRDC_FS2103_Ramularia_03.pdf
Pathogen
A podridão radicular de Rhizoctonia no trigo é causada por Rhizoctonia solani. O agente patogénico pode sobreviver durante muitos anos no solo sob a forma de esclerócios. Em alguns casos, também sobrevive como micélios em restos de plantas. Quando a temperatura aumenta, os esclerócios tornam-se activos e geram uma massa de hifas, permitindo a infeção ao fixarem-se às raízes. O micélio produz as chamadas "almofadas de infeção" e o fungo invade e coloniza o tecido da raiz. A infeção da raiz enfraquece as plântulas, resultando num crescimento deficiente, e reduz a capacidade de transporte de água e nutrientes para os tecidos da planta acima do solo, acabando por levar à morte da planta.
Sintomas
Numa fase inicial, aparecem manchas nuas, com tamanhos que variam de alguns centímetros a vários metros de diâmetro. Estas manchas nuas estão frequentemente associadas a plantas atrofiadas que podem apresentar amarelecimento, murchidão ou mesmo uma coloração púrpura. As plantas infectadas podem ser gravemente atrofiadas e mostrar sinais de stress hídrico ou deficiência de nutrientes, o que pode levar à morte prematura em alguns casos.
As lesões castanho-avermelhadas desenvolvem-se nos caules e nas raízes abaixo da linha do solo, ficando afundadas à medida que a doença progride. A podridão radicular pode reduzir a nodulação e fazer com que o córtex da raiz seja facilmente quebrável, resultando em pontas de lança castanhas.
Modelo FieldClimate
Modelo de risco de Rhizoctonia solani
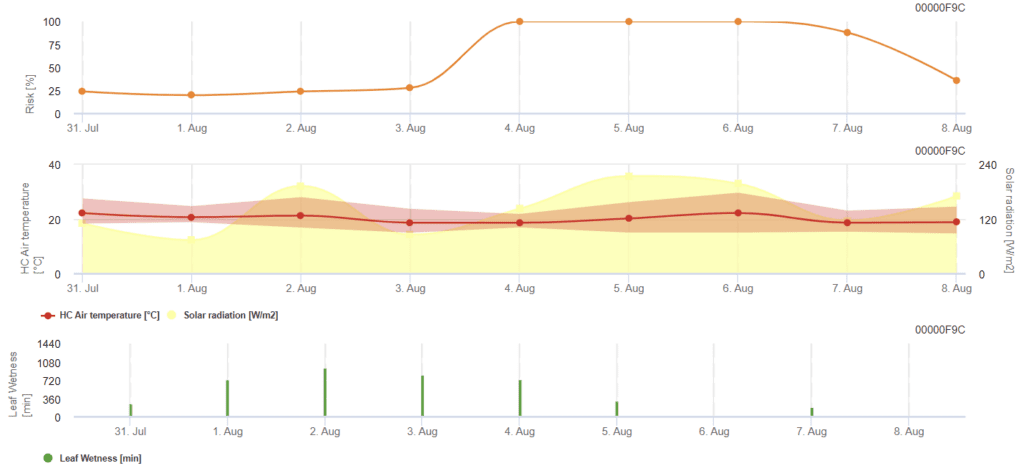
Sensores necessários:
- Temperatura do ar
- Radiação solar
- Humidade das folhas
O modelo calcula os períodos de risco de Rhizoctonia solani através da verificação das condições das últimas 120 horas. O resultado é um valor de risco de 0 a 100, indicando condições favoráveis para a doença.
Não são necessárias pulverizações quando o risco é baixo. Em períodos de risco moderado, o intervalo de pulverização pode ser prolongado e, em períodos de risco elevado, pode ter de ser reduzido ou pode ser recomendada a utilização de compostos mais eficazes.
Em caso de humidade consecutiva das folhas, acumula valores dependentes da temperatura para cada minuto:
- 12 °C a 15 °C: acumulação de 1 por minuto
- 16 °C a 17 °C: acumulação de 2 por minuto
- 18°C e mais: acumulação de 4 por minuto
No final dos períodos de humidade das folhas, avalia os valores acumulados:
- Valores > 4096: O risco aumenta em 64 pontos e a subtração de 4096 do valor
- Valores de resíduos > 2048: O risco aumenta em 16 pontos e a subtração de 2048 do valor
- Valores de resíduos >1024: O risco aumenta em 4 e a subtração de 1024 do valor
Se a radiação global for consecutivamente superior a 800 W/m², acumula o tempo em minutos e avalia os valores quando a radiação diminui:
- Valor > 512: Risco - 32 pontos, valor - 512
- Valor > 256: Risco - 8 pontos, valor - 256
- Valor > 128: Risco - 2 pontos, valor - 128
Literatura
- https://cropprotectionnetwork.org/encyclopedia/rhizoctonia-root-rot-of-wheat#:~:text=It%20is%20caused%20by%20Rhizoctonia,result%20in%20premature%20plant%20death.
- https://ahdb.org.uk/knowledge-library/rhizoctonia-stunt-symptoms-and-risk-in-cereals
- https://cropwatch.unl.edu/rhizoctonia-root-rot#:~:text=Disease%20Symptoms&text=These%20reddish%20brown%20lesions%20may,uneven%20because%20of%20stunted%20plants.
- https://extensionaus.com.au/FieldCropDiseasesVic/docs/identification-management-of-field-crop-diseases-in-victoria/soil-borne-diseases/rhizoctonia-root-rot/
- https://www.florimond-desprez.com/es/wp-content/uploads/sites/6/2015/11/rhizoctonia_eng.pdf
Pathogen
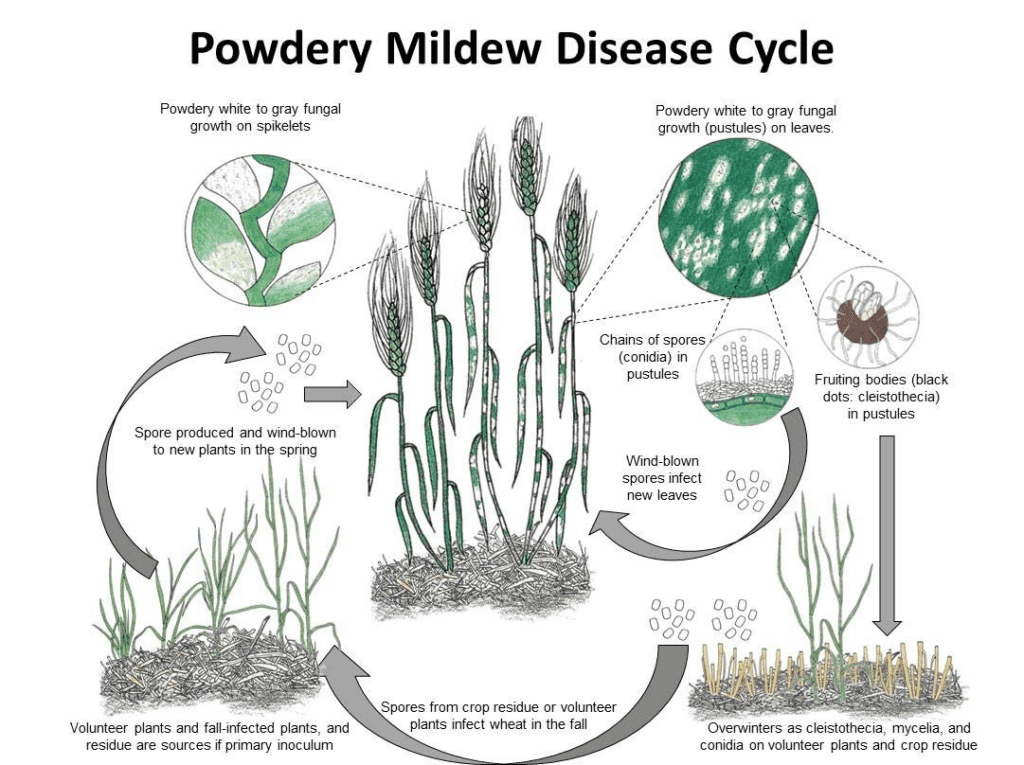
Blumeria graminis é o agente patogénico do oídio do trigo e da cevada. Pode hibernar sob a forma de micélios ou cleistotecas. Na primavera, os esporos germinam e infectam as plantas hospedeiras em condições frescas e húmidas, sem necessidade de água livre nas superfícies das plantas.
O fungo reproduz-se tanto assexuadamente como sexuadamente.
No ciclo assexuado, o agente patogénico passa por ciclos rápidos de infeção e de produção de conídios (esporos assexuados), que são fundamentais para a propagação da doença. Os conídios são produzidos no micélio a cada 7 a 10 dias em condições favoráveis. Uma vez dispersos pelo vento, estes conídios pousam num novo hospedeiro, germinam e formam apressórios e haustórios para extrair nutrientes da planta. Os conídios de Blumeria graminis podem germinar mesmo a baixa humidade e a várias temperaturas.
No ciclo sexual, o fungo produz cleistotecas, estruturas resistentes que podem sobreviver bem a condições adversas. As cleistotecas contêm ascósporos (esporos sexuais) e novas infecções são iniciadas quando estes esporos são libertados.
Sintomas

O oídio do trigo e da cevada afecta o rendimento do grão, reduzindo o número de cabeças e diminuindo o tamanho e o peso do grão. Quanto mais cedo ocorrer a infeção, maiores serão os danos.
A cevada apresenta uma resistência ao oídio relacionada com a idade. À medida que a cevada amadurece, em vez das manchas típicas de micélio, podem aparecer nas folhas manchas pretas e castanhas bem definidas ("tar-spots").
O trigo é particularmente suscetível ao oídio entre o período de perfilhamento e a maturação do leite, especialmente quando as folhas bandeira e as cascas são afectadas. No entanto, em comparação com a cevada, o trigo tolera melhor as infecções precoces.
O oídio branco-acinzentado desenvolve-se nas folhas, caules e cabeças. O crescimento do fungo é mais proeminente na superfície superior da folha, embora também se possa desenvolver na parte inferior. Inicialmente, as pústulas parecem brancas, mas gradualmente tornam-se cinzentas a castanhas à medida que amadurecem e podem coalescer, formando massas maiores, muitas vezes rodeadas de clorose. No lado oposto das folhas infectadas, os tecidos tornam-se amarelos, passando depois a castanhos ou bronzeados. Cleistothecia também são visíveis, como pequenos pontos pretos e redondos dentro de colónias cinzentas mais antigas. Em casos graves, as folhas podem morrer.
Modelo FieldClimate
Modelo de oídio do trigo
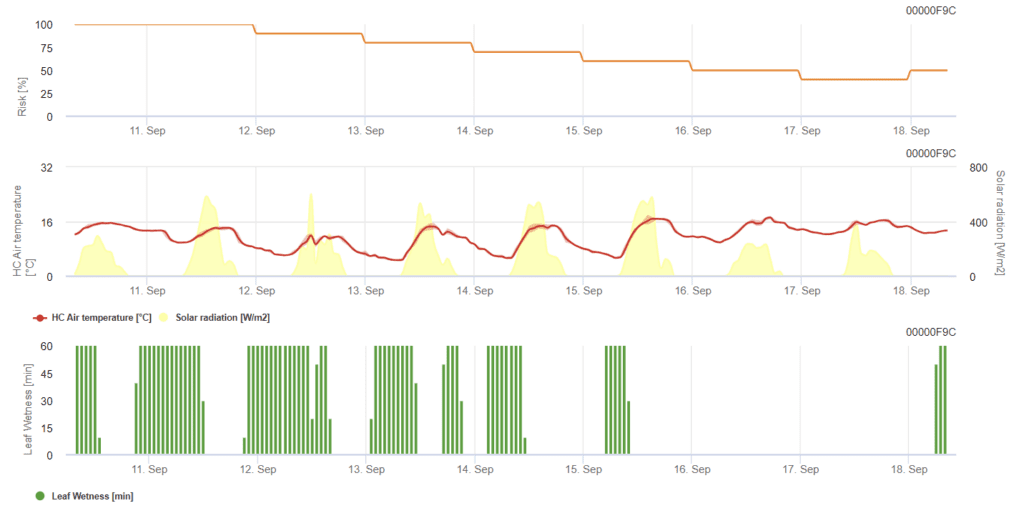
Sensores necessários:
- Temperatura do ar
- Humidade das folhas
- Radiação solar
O modelo determina o risco de oídio, variando numa escala de 0 (sem infeção) a 100% (infeção total). É relevante durante o estádio BBCH de 21 a 39 para a cevada e de 21 a 75 para o trigo e a aveia.
O estádio BBCH é uma escala de crescimento para descrever o desenvolvimento das plantas. Há dois dígitos, a escala varia de 00 a 99 - 00 refere-se ao tratamento de sementes e 99 é quando ocorre o tratamento pós-colheita. O primeiro dígito indica o estádio específico de desenvolvimento; 0 é a germinação, 1 é o desenvolvimento da folha, 2 é o perfilhamento, 3 é o alongamento do caule, 4 é o arranque, 5 é o aparecimento da inflorescência e a cabeça, 6 é a floração e a antese, 7 é o desenvolvimento do fruto, 8 é o amadurecimento e 9 é a senescência.
- BBCH 21: Perfilhamento - Início do perfilhamento; primeiro perfilho detetável.
- BBCH 39: Alongamento do caule - fase de folha bandeira; folha bandeira completamente desenrolada e lígula apenas visível.
- BBCH 75: Desenvolvimento do fruto - leite médio; teor de grãos leitosos e grãos que atingem o tamanho final, mas ainda verdes.
O risco aumenta com temperaturas entre 12 °C e 21 °C durante a maior parte do dia e com baixa radiação global. Em contrapartida, a humidade das folhas, a radiação global elevada e as temperaturas elevadas superiores a 32°C diminuem o risco.
Literatura
- Both, M., & Spanu, P. D. (2004). Blumeria graminis f. sp. hordei, um patógeno obrigatório da cevada. Revisões anuais de plantas, 11, 202-218.
- Cunfer, B. M. (2002). Oídio. Trigo para panificação: melhoramento e produção, 30, 317-330.
- Der Gräser, E. M. Krankheiten und Schädlinge des Getreides. Lancashire, P. D., Bleiholder, H., Boom, T. V. D., Langelüddeke, P., Stauss, R., Weber, E., & Witzenberger, A. (1991). Um código decimal uniforme para estádios de crescimento de culturas e infestantes. Anais de Biologia Aplicada, 119(3), 561-601. https://ohioline.osu.edu/factsheet/plpath-cer-11
Pathogen
Existem três ferrugens e agentes patogénicos importantes:
- Ferrugem da folha/ferrugem castanha: Puccinia triticina
- Haste/ Ferrugem preta: Puccinia graminis
- Risca/ferrugem amarela: Puccinia striiformis
A ferrugem castanha é a doença mais comum da ferrugem do trigo. A ferrugem negra é também conhecida como ferrugem de verão devido ao desenvolvimento abundante de teliosporos pretos brilhantes. É a doença mais devastadora da ferrugem, causando 50% das perdas num mês, em condições favoráveis. A ferrugem amarela é uma doença do trigo em climas mais frios, geralmente associada a altitudes mais elevadas e latitudes setentrionais. As suas caraterísticas são os urediniósporos de cor amarela. Também pode causar danos tão significativos como a ferrugem negra em casos graves.
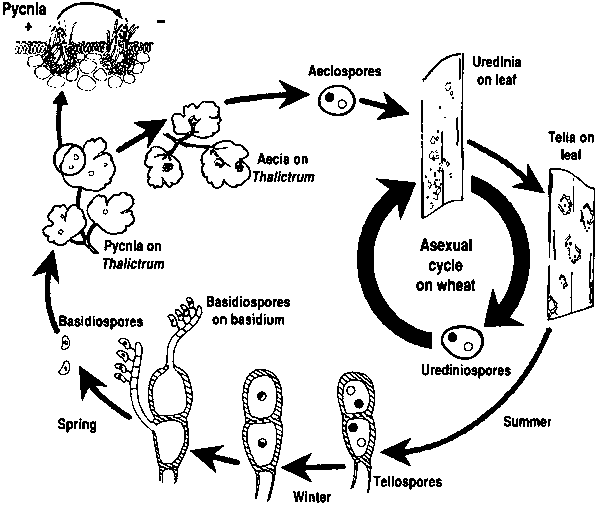
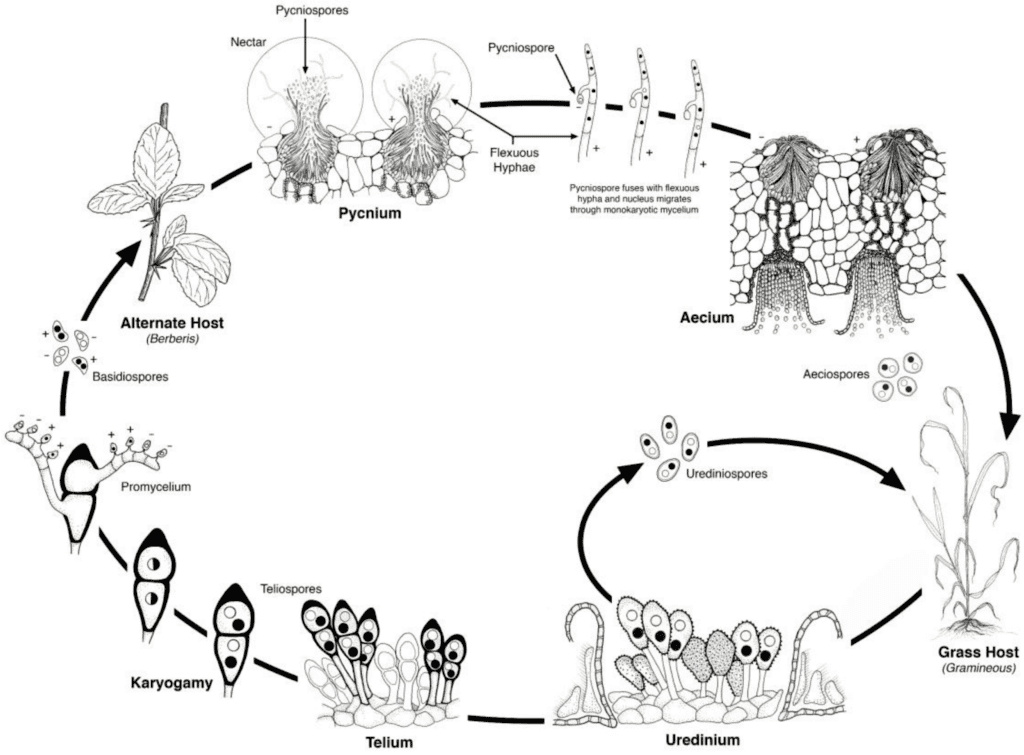
Os agentes patogénicos da ferrugem do trigo passam por ciclos de reprodução assexuada e sexual e necessitam de duas plantas hospedeiras - um hospedeiro económico e um hospedeiro alternativo. O hospedeiro económico é o trigo, e o hospedeiro alternativo é tipicamente uma planta daninha ou nativa, permitindo que os fungos passem pela fase sexual. Puccinia striiformis não é conhecido por ter qualquer hospedeiro alternativo, consistindo em fases urediniais e teliais.
No ciclo assexuado, os urediniósporos são produzidos na planta hospedeira e podem continuar a infectá-la repetidamente, levando a uma infeção rápida e generalizada em condições favoráveis.
O ciclo sexual do agente patogénico começa quando os teliosporos, formados em infecções urediniais na planta hospedeira, germinam e sofrem meiose para produzir basidiósporos. Estes basidiósporos são dispersos para um hospedeiro alternativo, onde iniciam a infeção, resultando na formação de picnias que contêm picniosporos e hifas receptivas. A fertilização ocorre quando os picniosporos de um tipo de acasalamento são transferidos para o oposto através do néctar. Após a fertilização, desenvolvem-se taças aeciais na parte inferior das folhas do hospedeiro alternativo, que libertam aeciosporos. Estes aeciosporos são dispersos e infectam novos hospedeiros, seguindo-se a infeção uredinial, completando o ciclo de vida do agente patogénico.
Sintomas

A) Puccinia graminis
B) Puccinia striiformis
C) Puccinia triticina
Puccinia triticina
P. triticina infecta principalmente as lâminas foliares, mas pode ocasionalmente afetar as bainhas das folhas, as glumas e os rebentos, formando pequenas manchas amarelas na superfície superior da folha. Estas manchas transformam-se em pústulas de cor laranja rodeadas por uma auréola amarela à medida que a doença progride. As pústulas produzem esporos alaranjados ou pretos visíveis nas folhas. Produz menos teliosporos, ao contrário da ferrugem negra. Algumas plantas podem apresentar manchas de hipersensibilidade ou uredinia rodeadas por áreas cloróticas ou necróticas.
Puccinia graminis
P. graminis A doença atinge principalmente os caules, mas também pode infetar folhas, bainhas, glumas, rebentos e até sementes. Inicialmente, surgem lesões castanho-avermelhadas e, à medida que a doença progride, formam-se teliosporos negros no interior das pústulas. Massas de urediniósporos vermelho-tijolo desenvolvem-se primeiro nos caules e nas bainhas das folhas e surgem pequenas manchas cloróticas, que se desenvolvem em pústulas lineares ou em forma de diamante que podem aumentar de tamanho. Quando atingem a maturidade, a produção de urediniósporos pára e é produzida uma camada de teliósporos pretos, fazendo com que os caules apareçam enegrecidos no final da estação. Em casos graves, a infeção enfraquece os caules das plantas, provocando o seu acamamento.
Puccinia striiformis
A caraterística do P. striiformis é a presença de pústulas amarelas de lados rectos. As TJey aparecem em faixas estreitas e alongadas de vários comprimentos, muitas vezes acompanhadas de clorose e necrose. À medida que a doença amadurece, os esporos amarelo-alaranjados desenvolvem-se no interior das pústulas e os tecidos circundantes tornam-se castanhos e secam, criando um aspeto queimado.
Modelo FieldClimate
Modelo da ferrugem da puccinia
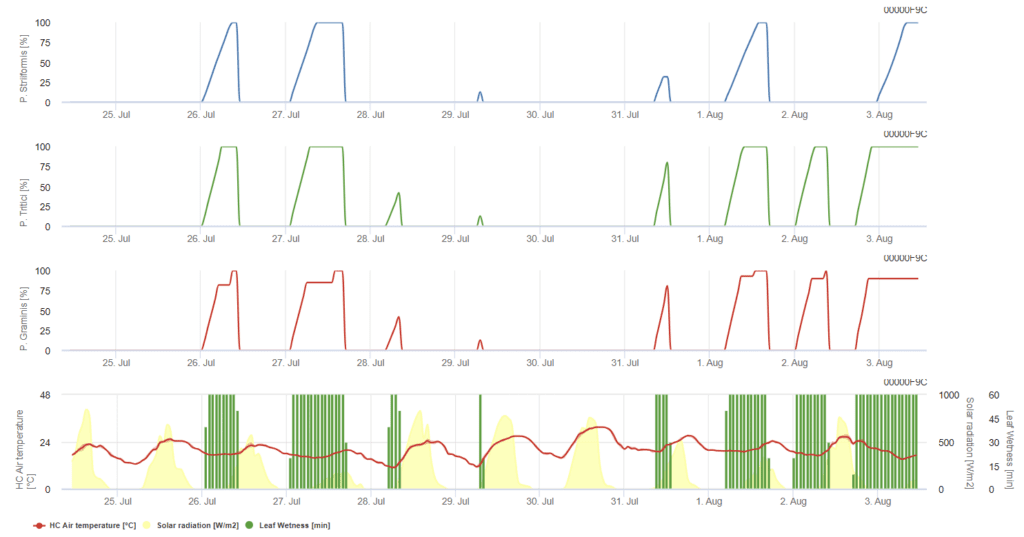
P. triticina, P. graminis, P. striiformis são apresentados em três gráficos de cores diferentes.
Modelo de infeção por Puccinia triticna
O modelo pressupõe que a infeção necessita de uma temperatura horária acumulada de 90°C de humidade foliar num intervalo de temperatura do ar de 5°C a 30°C. As condições são semelhantes às de P. graminismas com um limiar de temperatura inferior a 5°C.
Se for detectada uma infeção de 100%, deve ser considerada uma medida fitossanitária curativa. Se o risco for de 80% e as previsões meteorológicas apontarem para mais períodos de humidade foliar, podem ser efectuadas aplicações foliares de proteção.
Humidade das folhas para temperaturas médias horárias acumuladas de 90°C:
- (se T <= 22,5°C então ∑(Th) else ∑ (22,5-(Th-22,5))
- 5°C < Temp. < 30°C
Modelo de infeção por Puccinia graminis
As infecções ocorrem após algumas horas de humidade das folhas em condições de temperatura óptimas. O modelo pressupõe que a infeção necessita de uma temperatura horária acumulada de 80°C de humidade foliar numa gama de temperaturas do ar entre 10°C e 35°C. Prefere uma temperatura um pouco mais elevada do que P. triticina e a infecção tem de ser seguida pela luz solar.
Quando se verifica a infeção pelo 100%, as condições são favoráveis ao fungo, pelo que devem ser consideradas medidas de proteção das plantas.
Humidade das folhas para temperaturas médias horárias acumuladas de 80°C, seguida de um período de luz (150 W/m²) para temperaturas médias horárias acumuladas de 30°C:
- (se T <= 24°C então ∑(Th) senão ∑ (Th-24))
- 10°C < Temp. < 35°C
Modelo de infeção por Puccinia striiformis
Puccinia striiformis é a ferrugem do trigo dos climas frios, tendo a sua temperatura óptima já a partir dos 15°C. As infecções ocorrem após algumas horas de humidade foliar em condições de temperatura óptimas. O modelo pressupõe que a infeção necessita de uma temperatura horária acumulada de 80°C de humidade foliar num intervalo de temperatura do ar de 5°C a 20°C. Não se registam infecções em períodos de baixa intensidade luminosa.
Antes de o gráfico atingir 100%, pode ser feita uma aplicação de proteção, ao passo que, depois de atingir esse valor, devem ser consideradas estratégias de proteção curativa (sistémica).
Humidade da folha e luz para temperaturas médias horárias acumuladas para 80°C:
- (se T <= 15°C então ∑(Th) senão ∑ (Th-15))
- 5°C < Temp. < 20°C
Literatura
- Bolton, M. D., Kolmer, J. A., & Garvin, D. F. (2008). Ferrugem da folha do trigo causada por Puccinia triticina. Patologia molecular das plantas, 9(5), 563-575.
- Chen, X. M. (2005). Epidemiologia e controlo da ferrugem da risca [Puccinia striiformis f. sp. tritici] no trigo. Revista canadiana de patologia vegetal, 27(3), 314-337.
- Figueroa, M., Hammond-Kosack, K. E., & Solomon, P. S. (2018). Uma revisão das doenças do trigo - uma perspetiva de campo. Patologia molecular das plantas, 19(6), 1523-1536.
- Guide, A. Leaf, Stem, and Stripe Rust Diseases of Wheat. Leonard, K. J., & Szabo, L. J. (2005). Ferrugem do caule de pequenos grãos e gramíneas causada por Puccinia graminis. Patologia molecular das plantas, 6(2), 99-111.
- Kolmer, J. (2013). Ferrugem da folha do trigo: biologia do patógeno, variação e resistência do hospedeiro. Florestas, 4(1), 70-84.
- Singh, R. P., Huerta-Espino, J., Roelfs, A. P., & Curtis, B. C. (2002). As ferrugens do trigo. Crescimento, 2(25), 35.