

Agente patogeno
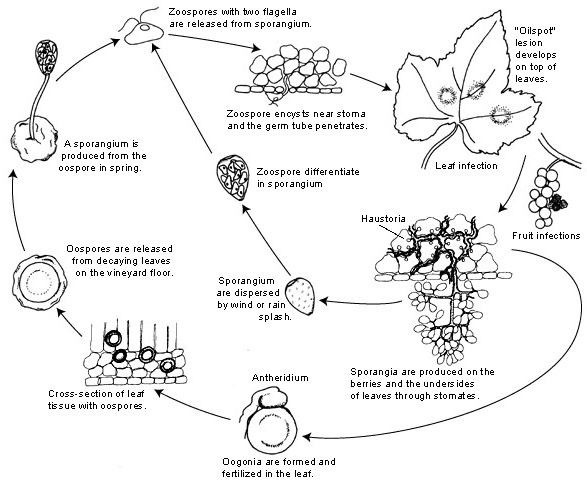
L'agente patogeno della peronospora della vite è Plasmopara viticola. È un parassita obbligato, il che significa che per crescere ha bisogno di organi verdi e freschi della vite. Presenta stadi riproduttivi sia asessuati che sessuati -zoospore e oospore. Esistono quindi due stadi di infezione: l'infezione primaria e quella secondaria.
L'infezione primaria inizia con la germinazione delle oospore svernanti, che formano gli sporangi in primavera, quando il clima diventa caldo. Gli sporangi vengono prodotti di notte, poiché la luce solare inibisce la sporulazione. Le oospore a parete spessa aiutano il patogeno a sopravvivere alle rigide condizioni invernali e ad avviare l'infezione primaria. Gli sporangi sono dispersi dalla pioggia o dal vento e rilasciano le zoospore nell'acqua libera. Per il successo dell'infezione è necessario che le foglie siano bagnate a sufficienza o che le precipitazioni siano sufficientemente lunghe.
L'infezione secondaria è possibile solo in presenza di macchie d'olio mature e avviene tramite zoospore e sporangi. Le zoospore e gli sporangi sono molto sensibili alla scarsa umidità e alla luce, che ne riducono la vitalità, il che significa che la maggior parte delle infezioni avviene subito dopo il rilascio. Le zoospore penetrano nei tessuti vegetali attraverso i tubi germinali e sviluppano ife per ulteriori infezioni. Queste ife formano lesioni oleose dopo un periodo di incubazione che varia da 5 a 21 giorni a seconda della temperatura - più bassa è la temperatura, più tempo ci vuole.
Quando il clima torna a essere freddo, sfavorevole per il patogeno, questo passa alla produzione sessuale, producendo oospore. Queste oospore rimangono dormienti durante l'inverno e il ciclo si ripete.
Sintomi

Sulla superficie superiore delle foglie compaiono piccole lesioni gialle, talvolta circondate da aloni brunastri. Le macchie oleose si espandono e si integrano con la maturazione e gli aloni svaniscono. Gli sporangi producono in seguito una muffa bianco-cotonica sulla parte inferiore delle lesioni, cui segue la necrosi. Le aree oleose marroni compaiono anche su germogli, steli e bacche. Nelle notti calde e umide, possono ricoprirsi di lanugine bianca a causa della sporulazione. Un'infezione grave causa la caduta prematura.
Modelli FieldClimate
- Nell'FieldClimate esistono due modelli: l'infezione primaria e quella secondaria.
- Il primo grafico mostra il tempo di incubazione dopo il completamento dell'infezione. I grafici inferiori mostrano l'andamento dell'infezione per un livello di gravità debole, moderato e grave. Si presume che ci sia un'infezione quando la curva crescente del grafico raggiunge 100% - l'irrorazione può essere presa in considerazione se si evidenzia un livello di infezione grave.
Infezione primaria
Sensori necessari:
- Temperatura dell'aria
- Umidità relativa
- Bagnatura delle foglie
- Precipitazioni
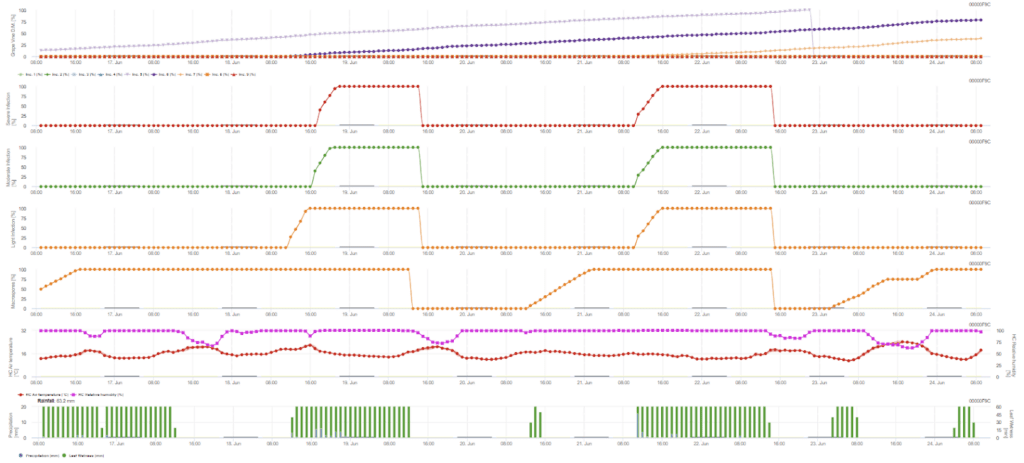
L'infezione primaria si verifica se il clima è adatto allo sviluppo degli sporangi. Ciò avviene finché le foglie sono bagnate o l'umidità relativa dopo la pioggia non scende al di sotto di 70%. Gli sporangi possono svilupparsi entro 16-24 ore, a seconda della temperatura. Una pioggia continua di 5 mm viene interpretata come una pioggia forte che può diffondere le zoospore.
Infezione secondaria
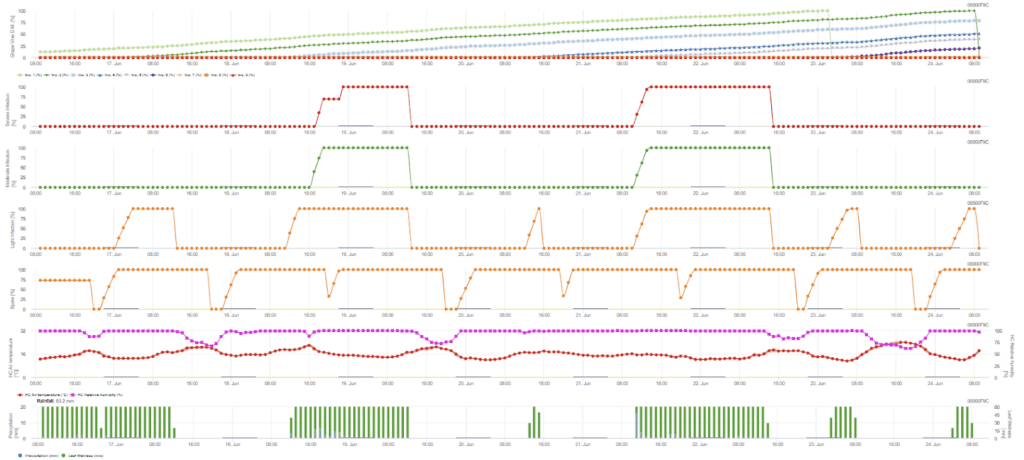
Sensori necessari:
- Temperatura dell'aria
- Umidità relativa
- Bagnatura delle foglie
- Precipitazioni
L'infezione secondaria si verifica se la temperatura è superiore a 12 °C e l'umidità relativa è superiore a 95%. Il tasso di produzione di sporangi aumenta con temperature fino a 23 °C. Se questa condizione si protrae per una temperatura oraria accumulata superiore a 50 °C, si presume che la sporulazione sia terminata e che esistano nuove sporangi nel vigneto. I 50 °C accumulati corrispondono a 4 ore con 13 °C o a 3 ore con 17 °C, per esempio. Gli sporangi muoiono rapidamente quando fa più caldo e più secco: quando l'umidità relativa scende al di sotto di 50%, la riportiamo a 0 e quando la temperatura supera i 29 °C, non può avvenire alcuna sporulazione.
Letteratura
- Ash, G. (2000). Peronospora della vite. L'Istruttore fitosanitario.
- Gessler, C., Pertot, I., & Perazzolli, M. (2011). Plasmopara viticola: una revisione delle conoscenze sulla peronospora della vite e sulla gestione efficace della malattia. Phytopathologia Mediterranea, 50(1), 3-44.
- Kennelly, M. M., Gadoury, D. M., Wilcox, W. F., Magarey, P. A., & Seem, R. C. (2007). Infezione primaria, produttività delle lesioni e sopravvivenza degli sporangi nel patogeno della peronospora della vite Plasmopara viticola. Phytopathology, 97(4), 512-522.
- Koledenkova, K., Esmaeel, Q., Jacquard, C., Nowak, J., Clément, C., & Ait Barka, E. (2022). Plasmopara viticola, agente causale della peronospora della vite: dalla tassonomia alla gestione della malattia. Frontiers in Microbiology, 13, 889472.
Agente patogeno
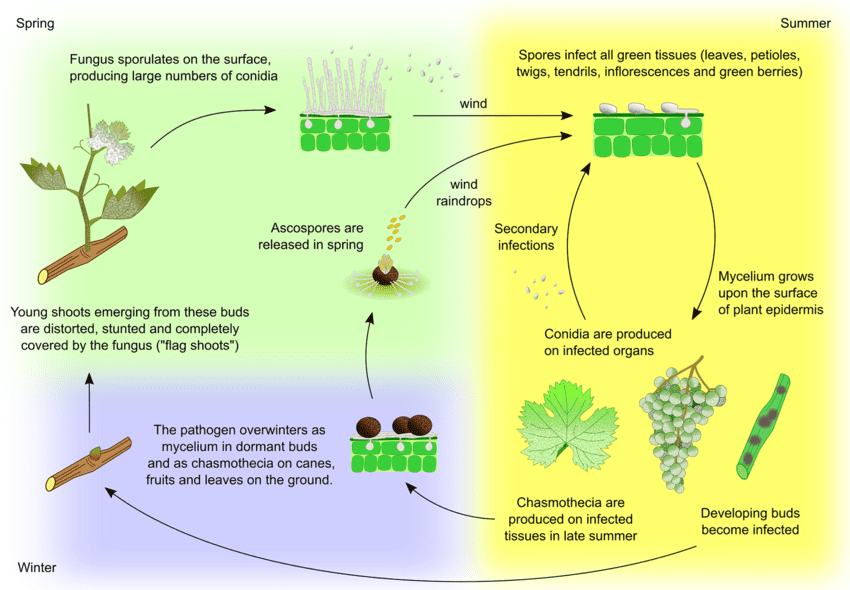
Il patogeno dell'oidio della vite è Uncinula necator. Le principali fonti di inoculo sono due: il micelio svernante e le ascospore dei cleistoteci, la cui importanza può variare a seconda della regione.
Il micelio provoca germogli a bandiera in primavera. I germogli a bandiera sono germogli parzialmente o completamente ricoperti di muffa che si formano da gemme infette latenti. Poiché all'interno si formano colonie, questi germogli sono ben visibili e spesso presentano foglie deformate.
I cleistoteci rilasciano le ascospore all'inizio della primavera. Le piogge sono un fattore importante per il rilascio delle ascospore. Con l'aumento della temperatura, la deiscenza avviene più frequentemente. Le ascospore germinano entro 12 ore, formando tubi germinali e appressori sui tessuti vegetali, con conseguente formazione di colonie sparse.
L'infezione secondaria avviene attraverso i conidi prodotti nelle colonie. Come le ascospore, germinano e sviluppano tubi germinali e appressori. U. necator non ha bisogno di acqua libera per l'infezione e di un'elevata umidità relativa per la formazione dei conidi. Quando le condizioni climatiche diventano meno favorevoli, il fungo forma cleistoteci. Sverna sopravvivendo nei cleistoteci o come micelio vegetativo nelle gemme infette dormienti e il ciclo si ripete.
Sintomi

I sintomi possono essere riscontrati su tutte le parti verdi della vite. I germogli sviluppano colonie e si ricoprono di micelio bianco-grigiastro, noto come "tralci bandiera". Producono foglie distorte. Le colonie di muffa si trovano anche sulle foglie, nella parte inferiore o su entrambi i lati. Gli acini sviluppano macchie grigio-cenere e, nei casi più gravi, si spaccano. Le canne presentano macchie rosso-marrone scuro e possono morire dalle punte.
Modelli FieldClimate
Infezione da ascospore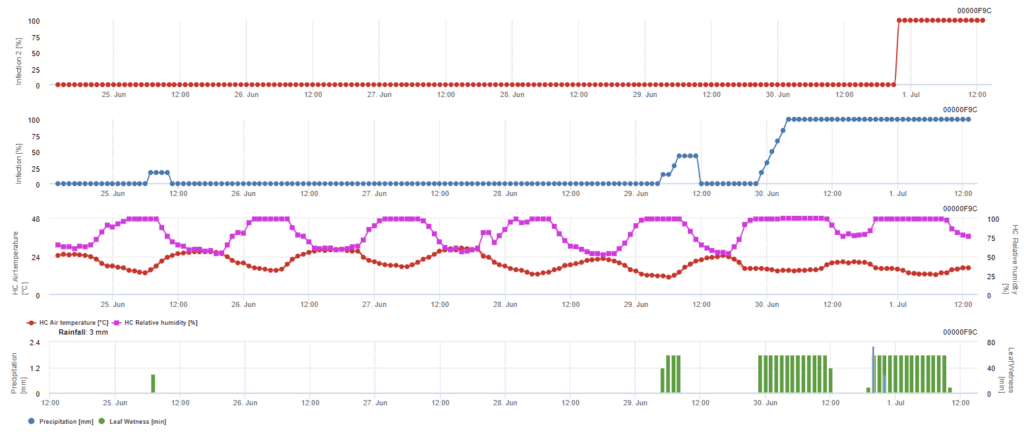
Sensori necessari:
- Temperatura dell'aria
- Umidità relativa
- Bagnatura delle foglie
- Precipitazioni
Questo modello è raccomandato quando le ascospore si formano durante il periodo invernale freddo (quindi non per l'infezione da parte del micelio). L'infezione da ascospore prevede il rilascio di ascospore e l'infezione iniziale in base alle temperature medie durante i periodi di bagnatura fogliare prolungata. Per il calcolo, sono necessari circa 2,5 mm di pioggia per rilasciare le ascospore, seguiti da un minimo di 8-12 ore di umidità fogliare e da temperature comprese tra 10 e 15°C. Una volta avvenuta l'infezione, il modello passa alla fase di valutazione del rischio di malattia (modello Californian Risk) e si basa sugli effetti della temperatura sul tasso di riproduzione del patogeno.
Modello di rischio californiano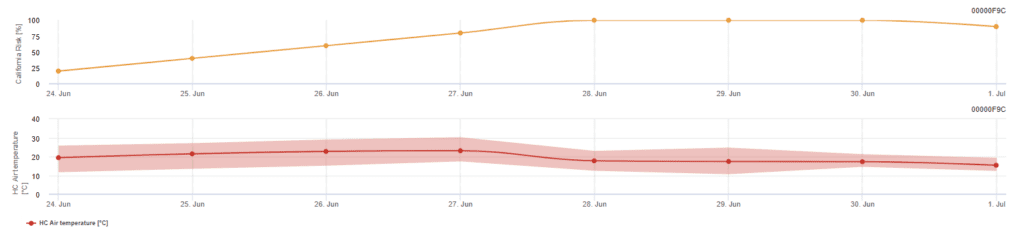
Sensori necessari:
- Temperatura dell'aria
Il modello si basa su studi di laboratorio in California. Dopo il rilascio delle ascospore e la germinazione (modello), il successivo sviluppo e la riproduzione dell'oidio sono influenzati dalle temperature. Il modello valuta le temperature e il rischio di sviluppo dell'oidio utilizzando un indice da 0 a 100 punti.
Per avviare l'indice di valutazione del rischio sono necessari tre giorni consecutivi con un minimo di sei ore consecutive di temperatura tra 21 e 30°C. Si guadagnano 20 punti per ogni giorno in cui si registrano 6 o più ore consecutive tra i 21 e i 32°C e si perdono 10 punti per quelli che non lo fanno o quando la temperatura supera i 32°C o scende sotto i 21°C.
Valori bassi dell'indice, compresi tra 0 e 30, indicano che il patogeno non si riproduce. Un indice di 40~50 è considerato moderato e implica un tasso di riproduzione dell'oidio di circa 15 giorni. Valori dell'indice superiori a 60 indicano che il patogeno si riproduce rapidamente (ogni 5 giorni) e che il rischio di un'epidemia è elevato.
Modello di rischio Pessl Instruments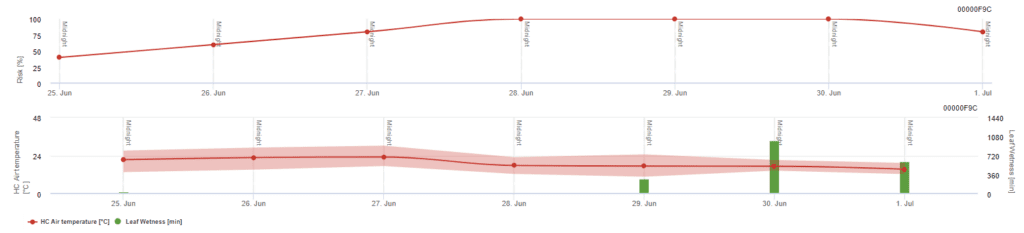
Sensori necessari:
- Temperatura dell'aria
- Bagnatura delle foglie
Oltre alla temperatura dell'aria, la bagnatura fogliare può essere un fattore guida, poiché lunghi periodi di bagnatura fogliare portano all'insediamento di un fungo antagonista (Ampelomyces quisqualis), che porta a una diminuzione di U. necator. I fondamenti del modello sono equivalenti a quelli del modello di rischio californiano, ma in questo modello viene considerata anche l'umidità fogliare. Una bagnatura fogliare superiore alle 8 ore comporta una diminuzione di 10 punti.
Valori bassi dell'indice, compresi tra 0 e 20, indicano che il patogeno non si sta riproducendo. Un indice di 20~60 è considerato moderato ed è valido un normale intervallo di irrorazione. Valori dell'indice superiori a 60 indicano che il patogeno si sta riproducendo rapidamente (ogni 5 giorni) e che il rischio di un'epidemia è elevato, per cui si raccomanda di ridurre l'intervallo di irrorazione.
Letteratura
- Gadoury, D. M., & Pearson, R. C. (1990). Deiscenza dell'ascocarpo e fuoriuscita delle ascospore in Uncinula necator. Fitopatologia, 80(4), 393-401.
- Gadoury, D. M., & Pearson, R. C. (1990). Germinazione delle ascospore e infezione di Vitis da parte di Uncinula necator. Fitopatologia, 80(11), 1198-1203.
- Hall, T. W. (2000). Epidemiologia dell'oidio dell'uva, Uncinula necator, nella Willamette Valley.
- Halleen, F. e Holz, G. (2001). Una panoramica sulla biologia, l'epidemiologia e il controllo di Uncinula Necator (oidio) sulla vite, con riferimento al Sudafrica. South african journal of Enology and Viticulture, 22(2), 111-121.
- Rügner, A., Rumbolz, J., Huber, B., Bleyer, G., Gisi, U., Kassemeyer, H. H., & Guggenheim, R. (2002). Formazione di strutture svernanti di Uncinula necator e colonizzazione della vite in condizioni di campo. Patologia vegetale, 51(3), 322-330.
Agente patogeno
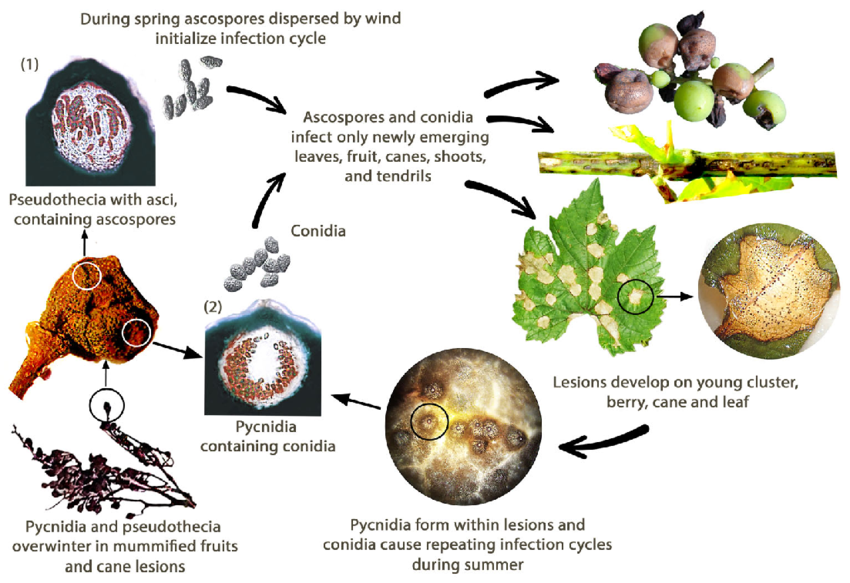
Il marciume nero della vite è causato dal fungo Guignardia bidwellii. Il fungo sverna in varie parti della vite e può sopravvivere per oltre due anni nelle lesioni dei tralci infetti.
In primavera, i picnidi producono conidi (spore asessuate) e gli pseudoteci generano ascospore (spore sessuate). I conidi si disperdono tipicamente su brevi distanze, mentre le ascospore possono infettare su distanze maggiori. Queste spore sono trasportate dal vento e dalla pioggia per infettare i tessuti giovani di ospiti sensibili, dando inizio a infezioni primarie. I conidi fungono anche da inoculo secondario, contribuendo alla rapida e ripetuta diffusione della malattia.
Durante il mese di agosto, i picnidi si trasformano in uno stadio svernante che produce pseudoteche, le quali generano ascospore. Queste ascospore sono importanti fonti per le infezioni primarie nella primavera successiva.
Sintomi

Si sviluppano macchie bruno-rossastre che alla fine si fondono, spesso circondate da piccoli punti neri, che sono strutture fungine note come picnidi che producono conidi. Le foglie giovani e in rapida crescita sono particolarmente sensibili a questa infezione.
Le infezioni dei frutti si verificano in genere dopo la caduta del calice, con la maggior parte dei sintomi che si manifestano quando il frutto è a metà o quasi a grandezza naturale. Inizialmente emergono piccole macchie, circondate da un anello marrone, che poi si allargano e si scuriscono, ricoprendo infine l'intera bacca man mano che la malattia si sviluppa. Nel giro di pochi giorni, le bacche infette possono mummificarsi, frantumarsi o cadere prematuramente.
Inoltre, vengono colpite anche altre parti della pianta, come germogli, steli e viticci. Si sviluppano lesioni di forma ovale di colore viola o nero, sulla cui superficie si spargono i picnidi.
Modello FieldClimate
Viticoltura Modello di marciume nero
Sensori necessari:
- Temperatura dell'aria
- Umidità relativa
- Bagnatura delle foglie
- Precipitazioni
Il modello è originariamente basato sulla letteratura pubblicata da Spotts, ma con la modifica di Daniel Molitor. La revisione ha introdotto tre classi di gravità (leggera, moderata e grave) simili a quelle dei modelli di ticchiolatura del melo. A temperature calde, il periodo di bagnatura fogliare di 8 ore è sufficiente a favorire le infezioni e si presume che un'infezione sia completata quando un grafico raggiunge 100%.
La gravità dell'infezione dipende dalla durata della temperatura ottimale e dal periodo di umidità del fungo. Le infezioni che soddisfano i criteri di Spotts sono classificate come infezioni leggere. Le infezioni che soddisfano i criteri Spott di 150% e 200% sono classificate come infezioni moderate e gravi, rispettivamente.
A seconda della pressione dell'infezione, le infezioni devono essere coperte in via preventiva o deve essere applicato uno spray curativo poco dopo l'infezione. Nel clima moderato e semi-arido della Mosella o della maggior parte delle zone vinicole austriache, non è consigliabile irrorare dopo aver accertato un'infezione leggera.
Letteratura
- Wilcox, Wayne F. "Marciume nero Guignardia bidwellii". Scheda di identificazione delle malattie n. 102GFSG-D4. 2003. Cornell. 24 ottobre 2010
- http://www.nysipm.cornell.edu/factsheets/grapes/diseases/grape_br.pdf
- Ellis, Michael A. "Scheda informativa sulle risorse agricole e naturali: Marciume nero della vite". Dipartimento di patologia vegetale. Università dell'Ohio State Extension. 2008
- http://ohioline.osu.edu/hyg-fact/3000/pdf/HYG_3004_08.pdf
- Molitor, D. (2009). Ricerche sulla biologia e sull'alimentazione della zigolo nero (Guignardia bidwellii) nel bosco. Gesellschaft zur Förderung der Forschungsanstalt Geisenheim.
- Ries, S. M. (1999). Rapporti sulle malattie delle piante: Marciume nero della vite. Gestione integrata dei parassiti presso l'Università dell'Illinois. http://ipm. illinois. edu/diseases/series700/rpd703.
- Spotts, R. A. (1977). Effetto della durata dell'umidità fogliare e della temperatura sull'infettività di Guignardia bidwellii sulle foglie d'uva. Phytopathology, 67(11), 1378-1381.
- Szabó, M., Csikász-Krizsics, A., Dula, T., Farkas, E., Roznik, D., Kozma, P., & Deák, T. (2023). Marciume nero dell'uva (Guignardia bidwellii) - Una panoramica completa. Horticulturae, 9(2), 130.
- http://extension.cropsciences.illinois.edu/fruitveg/pdfs/771-BlackRotOfGrape.pdf
- https://www.missouribotanicalgarden.org/gardens-gardening/your-garden/help-for-the-home-gardener/advice-tips-resources/insects-pests-and-problems/diseases/fruit-spots/black-rot-of-grapes
Agente patogeno

Lobesia botrana compie in genere due o tre generazioni all'anno, a seconda della regione. La prima generazione (maggio e giugno) colpisce solo i fiori, quindi il trattamento è necessario solo se le popolazioni di tignole sono particolarmente elevate. La seconda generazione (luglio e agosto) e la terza generazione causano i danni maggiori; la terza generazione è particolarmente dannosa perché coincide con la maturazione dell'uva. Nei climi più caldi può verificarsi una quarta generazione, ma in genere il trattamento non è necessario perché coincide con la vendemmia.
La falena sverna come pupa in bozzoli e, quando le temperature si alzano in primavera, emergono gli adulti della prima generazione, con i maschi che in genere compaiono prima delle femmine. Il volo della prima generazione inizia in prossimità della rottura delle gemme e dura da 4 a 5 settimane, durante le quali avviene l'accoppiamento. Dopo uno o due giorni dall'accoppiamento, le femmine depongono da 80 a 160 uova.
Le uova, del diametro di 0,6-0,8 mm, sono visibili a occhio nudo. Inizialmente di colore bianco crema, diventano gialle con lo sviluppo dell'embrione, con il colore nero della testa della larva. Le uova si schiudono dopo 66 gradi centigradi (DDC), circa 7-11 giorni. Le larve si attaccano alle parti del fiore e il loro sviluppo dura da 20 a 30 giorni. Segue l'accoppiamento e gli adulti emergono da 6 a 14 giorni dopo.
Il "grado-giorno" si calcola misurando quanto la temperatura media giornaliera supera una certa temperatura di base necessaria per lo sviluppo. Man mano che le temperature salgono al di sopra della soglia di base, i gradi-giorno si accumulano.
Gli adulti sono lunghi circa 6-8 mm e hanno un'apertura alare di 11-13 mm. Le femmine sono leggermente più grandi dei maschi, anche se entrambi hanno ali simili a mosaico.
Le uova della seconda e terza generazione si schiudono più rapidamente, entro 3-5 giorni. Le femmine depongono le uova singolarmente su bacche in ombra e, una volta schiuse, le larve entrano nelle bacche e le scavano. In autunno, le notti superiori alle 11 ore provocano la diapausa, una fase di riposo che consente alle pupe di tollerare meglio il freddo, permettendo loro di svernare.
Sintomi
I bruchi della prima generazione si nutrono delle infiorescenze della vite, scavando i boccioli dei fiori e facendoli ruotare per proteggerli, con conseguenti danni all'interno del grappolo. Nella seconda generazione, le larve prendono di mira gli acini in via di sviluppo, creando punti d'ingresso per patogeni fungini quali Botrytis cinerea. Questa alimentazione diretta provoca danni visibili, tipicamente contrassegnati da macchie scure intorno ai punti di alimentazione. La terza generazione causa i danni più significativi, poiché le larve penetrano e si nutrono delle bacche in fase di maturazione.
Rimangono bacche raggrinzite ed escrementi larvali. Brandelli di tessuto epidermico dell'acino rimangono attaccati ai pedicelli, insieme alla "pelle" dell'acino secca e scavata.
Modello FieldClimate
Tignola della vite Modello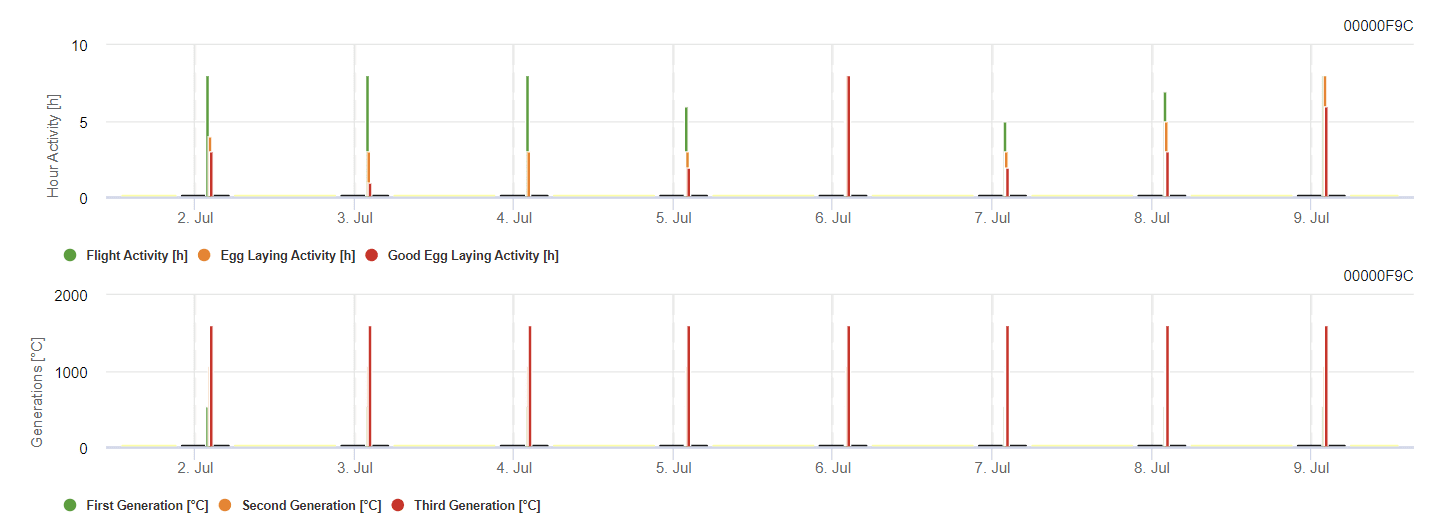
Sensore necessario:
- Temperatura dell'aria
Il modello calcola il rischio in base alla temperatura dell'aria. Il primo grafico mostra i periodi di attività di volo con deposizione di uova e buona attività di deposizione di uova per l'insetto. Il secondo grafico mostra la generazione attesa nel periodo. Nel terzo grafico vengono visualizzati i dati climatici, che mostrano l'accumulo di gradi giorno (temperature > 8°C fino a 24°C per ora divise per 24).
Letteratura
- Varela, L. G., Smith, R. J., Cooper, M. L., & Hoenisch, R. W. (2010). Tignola della vite, Lobesia botrana. Vigneti della Napa Valley. Pratica. Winery Vineyard, 2010, 1-5.
- https://ipm.ucanr.edu/invasive-and-exotic-pests/european-grapevine-moth/
- https://www.ages.at/en/plant/plant-health/pests-from-a-to-z/grape-berry-moth
Agente patogeno
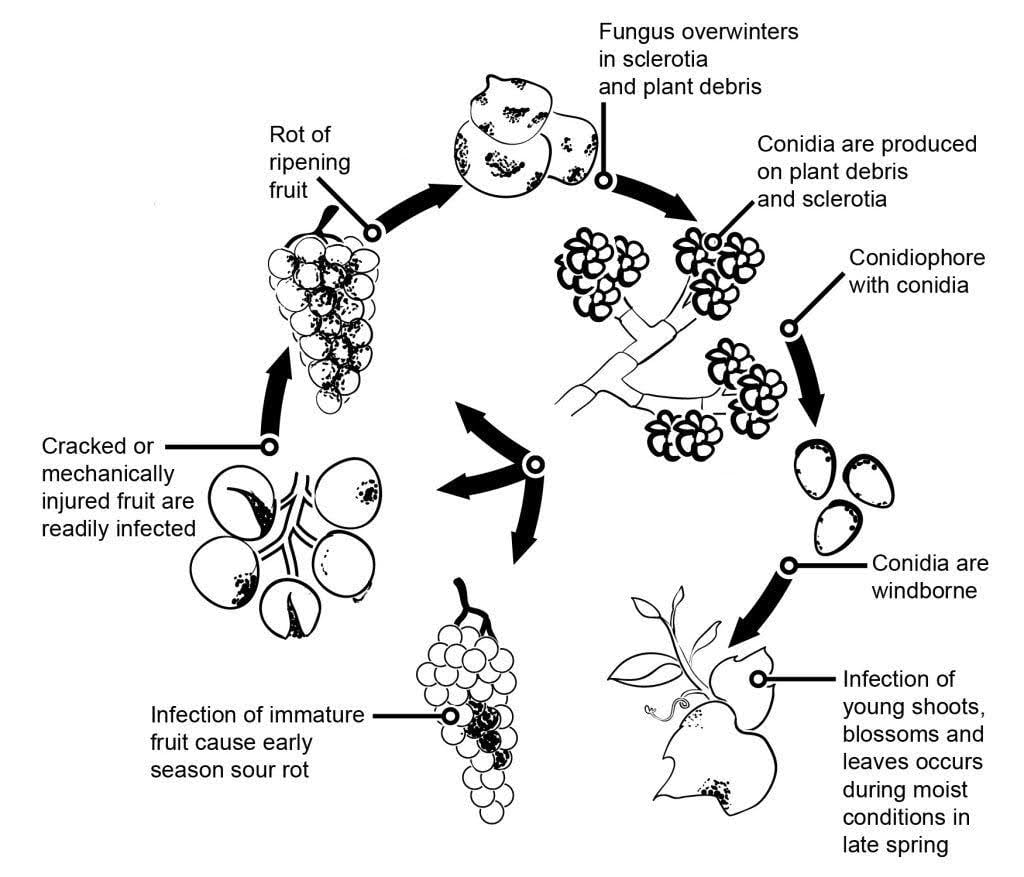
L'agente patogeno della muffa grigia nella vite è Botrytis cinerea.
L'epidemiologia della muffa grigia è significativamente influenzata da infezioni latenti iniziate da infezioni floreali. Sono stati identificati diversi percorsi da fiore a frutto: Il fungo infetta lo stilo del fiore e raggiunge l'ovulo, dove rimane latente grazie al meccanismo di difesa preformato della pianta. Anche le infezioni attraverso stami, petali e sepali sono fondamentali. B. cinerea può infettare gli stami e crescere sistematicamente verso il ricettacolo, diffondendosi al peduncolo e ai tessuti vascolari delle bacche. Altre vie prevedono la crescita saprofitica del patogeno.
I miceli o gli sclerozi svernanti sono le principali fonti di infezione in primavera. I conidi (spore asessuate) vengono prodotti e dispersi dal vento e dalla pioggia. Una volta atterrati, i conidi germinano, formando tubi germinali e appressori che penetrano e infettano la pianta. Il fungo può rimanere latente fino alla maturazione del frutto e all'aumento del contenuto di zucchero. I conidi generati dalle fonti primarie di inoculo seguono un ciclo diurno di iniziazione, produzione e diffusione.
Sebbene lo stadio sessuale di B. cinerea sia raramente osservato, il patogeno diventa più attivo con la maturazione delle bacche. L'aumento del contenuto di zuccheri nelle bacche aumenta la suscettibilità all'infezione. Allo stesso modo, i fiori sono altamente suscettibili quando si estinguono, e l'abbondanza di polline aumenta la gravità dell'infezione.
Sintomi

La muffa grigia infetta principalmente gli acini d'uva maturi, che inizialmente appaiono morbidi e impregnati d'acqua. Con il tempo, gli acini diventano bruno-rossastri e raggrinziscono. In condizioni favorevoli, si ricoprono di masse grigie di miceli e conidi fungini. Gli acini sani possono infettarsi anche per contatto diretto con quelli malati.
Il fungo può anche causare una peronospora dei fiori, che può portare a significative perdite di raccolto all'inizio della stagione. Sebbene le infezioni fogliari siano rare, quando si verificano iniziano come macchie verdi opache lungo le venature, che alla fine si sviluppano in lesioni necrotiche.
Modello FieldClimate
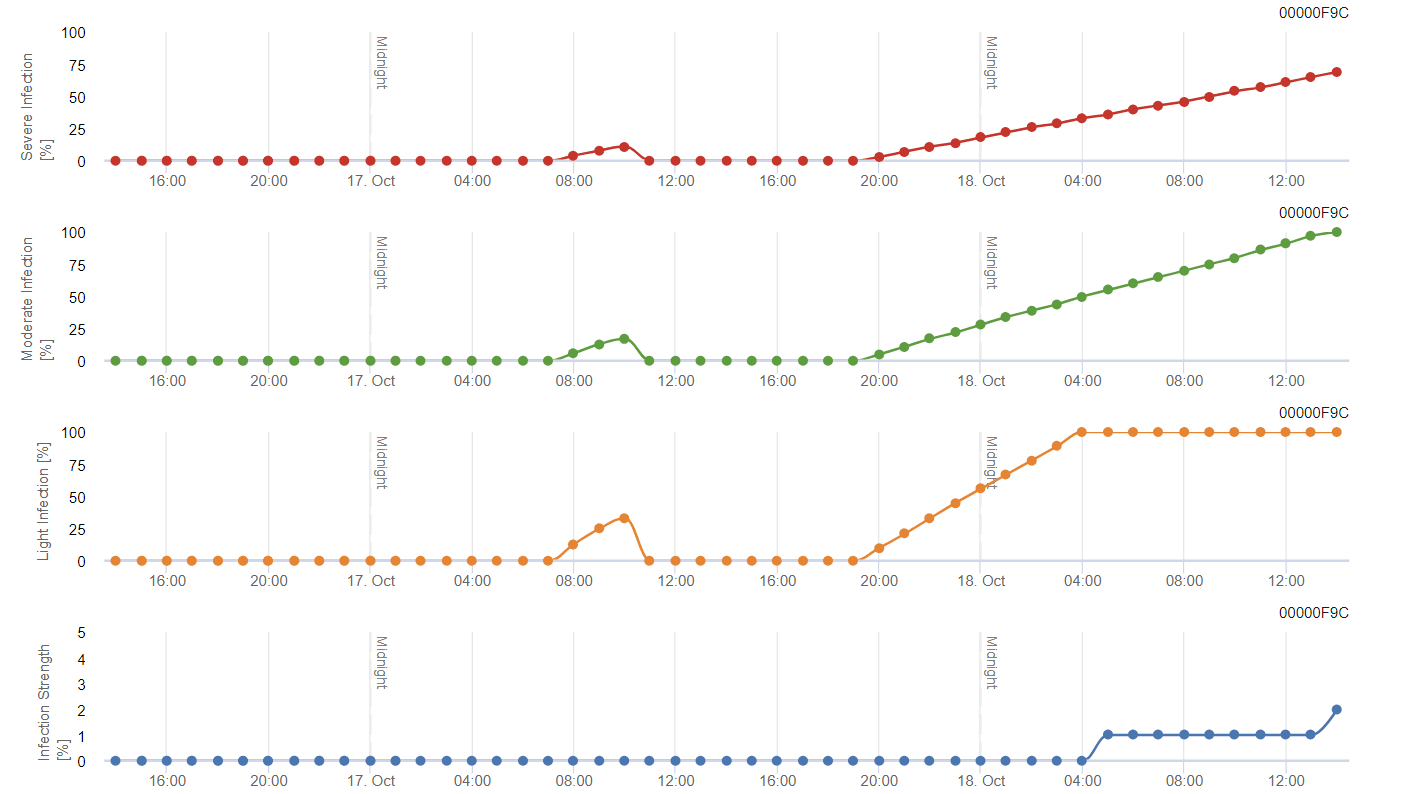
Modello di stampo grigio generale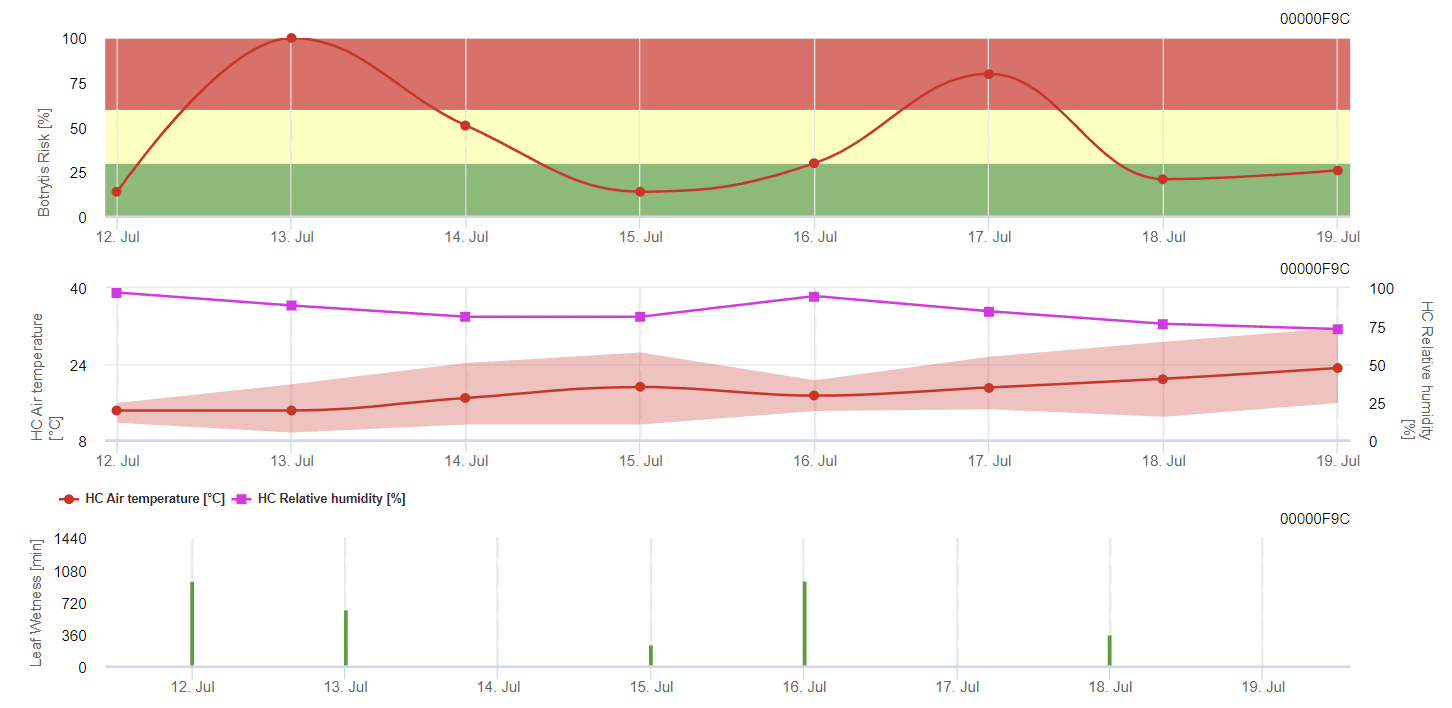
Sensori necessari:
- Temperatura dell'aria
- Umidità relativa
- Bagnatura delle foglie
B. cinerea è legato al clima umido. Il fungo richiede un'elevata umidità relativa o la presenza di acqua libera per l'infezione. I periodi caldi, con un prolungato periodo di bagnatura delle foglie, comportano un aumento del rischio di B. cinerea, mentre i periodi secchi lo riducono. L'infezione avviene sui giovani germogli durante i lunghi periodi umidi o le dannose tempeste di grandine.
Il modello calcola il rischio in un valore compreso tra 0 e 100%. Questo valore indica la pressione di B. cinerea al momento - se abbiamo un valore di 100%, significa che c'è stato più volte un periodo di bagnatura abbastanza lungo da infettare il tessuto suscettibile. Calcoliamo i cosiddetti "punti di bagnatura" tra la bagnatura delle foglie e la temperatura, con un massimo iniziale di 38400 punti (inizio della stagione, che presenta un rischio di 30%). Dopo questo periodo, ogni periodo di bagnatura con circa 4000 punti di bagnatura aumenta il rischio di 10%, o dall'altra parte, ogni periodo di siccità riduce il rischio di ⅕ del valore precedente.
Letteratura
- Broome, J. C., English, J. T., Marois, J. J., Latorre, B. A., & Aviles, J. C. (1995). Sviluppo di un modello di infezione per il marciume dei grappoli di Botrytis basato sulla durata dell'umidità e sulla temperatura. Fitopatologia, 85(1), 97-102.
- Elmer, P. A. e Michailides, T. J. (2007). Epidemiologia della Botrytis cinerea nei frutteti e nelle colture di vite. In Botrytis: biology, pathology and control (pp. 243-272). Dordrecht: Springer Netherlands.
- Williamson, B., Tudzynski, B., Tudzynski, P., & Van Kan, J. A. (2007). Botrytis cinerea: la causa della malattia della muffa grigia. Patologia vegetale molecolare, 8(5), 561-580.
- Ciliberti, N., Fermaud, M., Roudet, J., & Rossi, V. (2015). Le condizioni ambientali influenzano l'infezione da Botrytis cinerea degli acini d'uva maturi più del ceppo o del genotipo del trasposone. Fitopatologia, 105(8), 1090-1096.
Agente patogeno
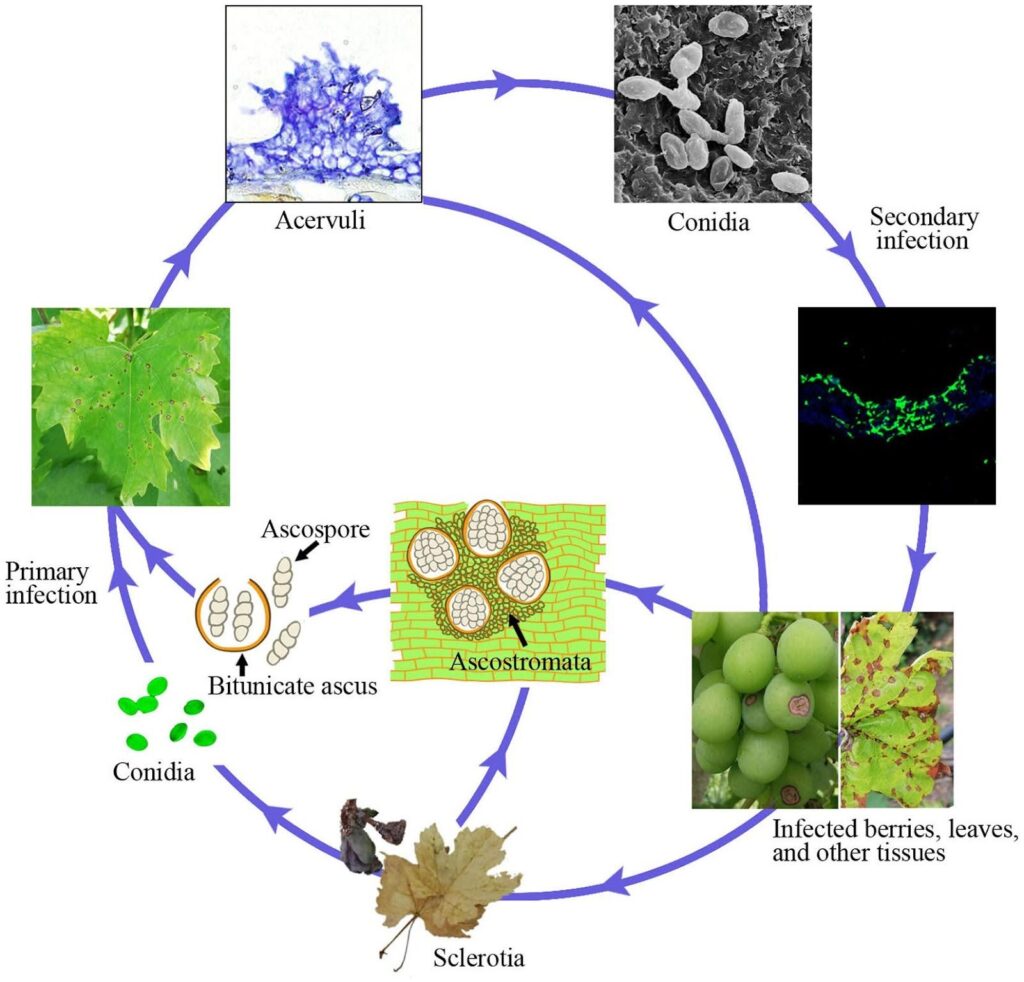
L'antracnosi della vite è causata dal patogeno Elsinoe ampelina.
Le canne infette sono la principale fonte della malattia. Gli sclerozi e i miceli che sopravvivono nelle lesioni e negli acini durante l'inverno si attivano in primavera e producono ascospore (spore sessuali) e conidi (spore asessuali) in condizioni di umidità (pioggia o rugiada per 24 ore) in un intervallo di temperatura compreso tra 2 e 40°C.
Queste spore vengono disperse in nuovi tessuti dagli spruzzi di pioggia e dal vento e, una volta atterrate, germinano e formano tubi germinali e appressori, dando inizio a nuove infezioni. Possono infettare nuove foglie, germogli, viticci e giovani bacche. Il clima caldo riduce la durata dell'umidità necessaria per l'infezione iniziale e il periodo di incubazione. La germinazione ottimale delle spore avviene a 25~30°C, con un minimo di 3~4 ore di bagnatura delle foglie. In caso di infezioni a una temperatura di circa 10°C, il periodo di incubazione è di circa 14 giorni.
Con il progredire della colonizzazione, emergono gli acervuli e vengono prodotti nuovi conidi, che fungono da inoculo per le infezioni secondarie. Sono responsabili di una rapida e ulteriore infezione nella stagione.
Sintomi
E. ampelina attacca le parti aeree e succulente della vite, compresi germogli, foglie, piccioli, viticci, rachide e acini; le lesioni più comuni sono quelle sui germogli e sugli acini. La vite presenta una resistenza ontogenetica al fungo, per cui i tessuti giovani sono più suscettibili.
All'inizio compaiono piccole macchie bruno-rossastre che si ingrandiscono con il tempo umido, diventando leggermente infossate con un centro grigio circondato da un margine scuro. A volte i sintomi possono assomigliare a lesioni da grandine. Il centro necrotico delle foglie di solito cade, creando un aspetto "a buchi".
In caso di infezione grave, le piante possono presentare defogliazione precoce, crescita stentata delle canne, rottura del fusto, caduta degli acini e ritardo nello sviluppo e nella maturazione delle bacche.
Modello FieldClimate
Modello di antracnosi viticola
Sensori necessari:
- Temperatura dell'aria
- Umidità relativa
- Bagnatura delle foglie
- Precipitazioni
Il modello calcola il rischio di antracnosi considerando la temperatura dell'aria, l'umidità relativa, la bagnatura delle foglie e le precipitazioni.
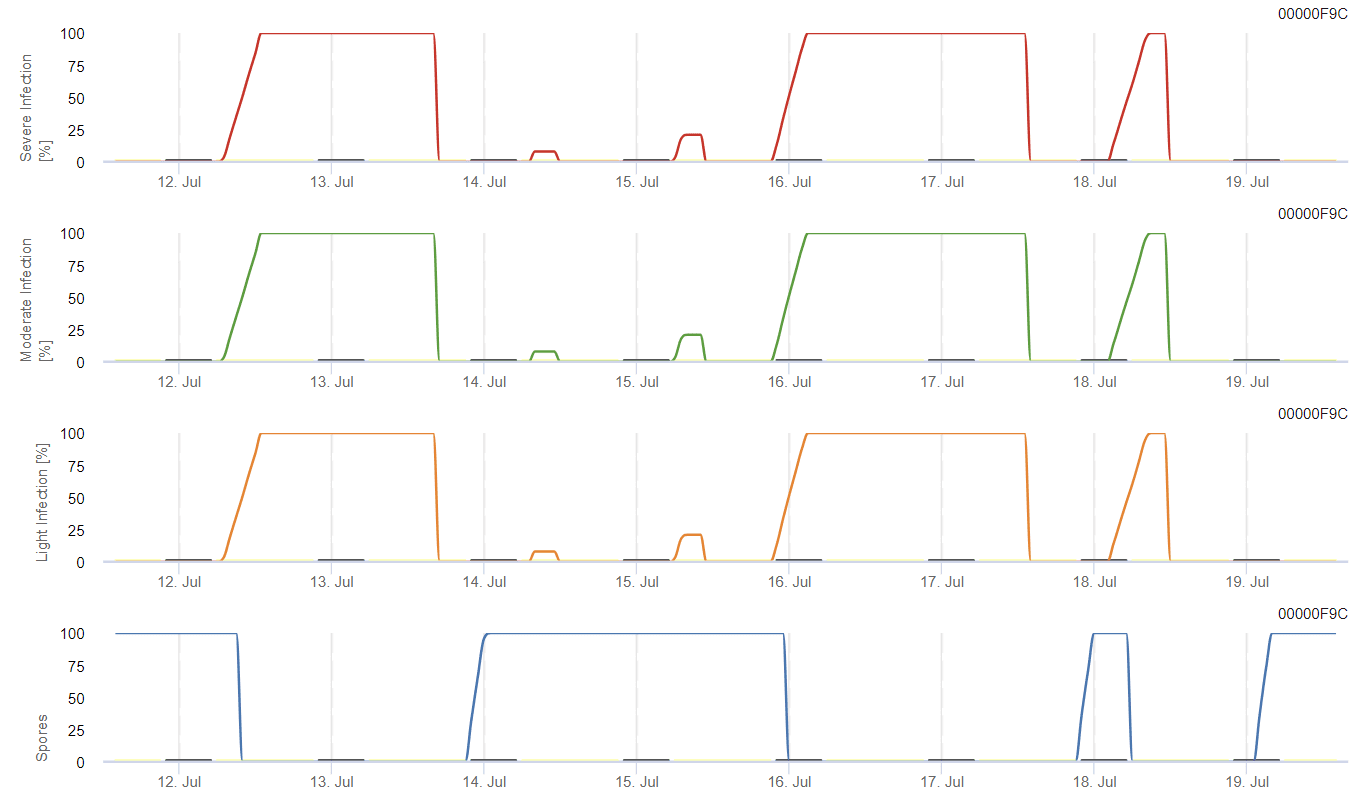
Il possibile sviluppo di un'infezione debole, moderata e grave viene visualizzato in grafici separati. Si presume che l'infezione sia completa quando la curva crescente raggiunge 100%. L'ultimo grafico mostra la sporulazione di E. ampelinaSe il valore raggiunge 100%, si presume che le spore di E. ampelina sono presenti.
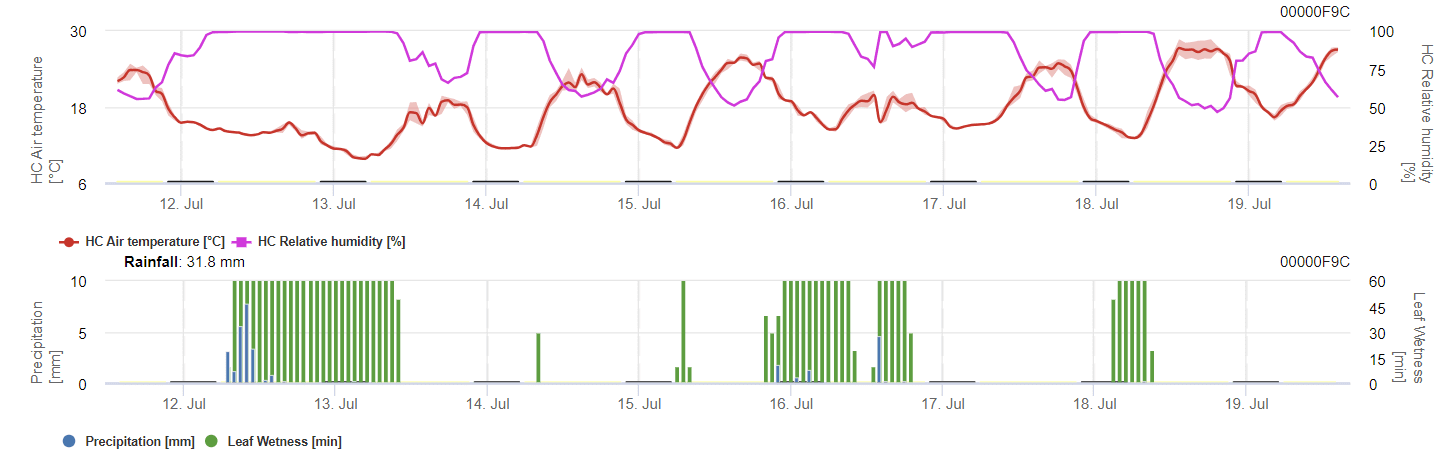
Affinché le spore svernanti si sviluppino, le condizioni devono rispettare un intervallo di temperatura compreso tra 2 e 40°C, un'umidità relativa superiore a 90% o l'umidità delle foglie. Una volta che lo sviluppo delle spore raggiunge 100%, l'infezione inizia a essere calcolata. La gravità dell'infezione dipende dalle condizioni di umidità (eventi di pioggia). Tuttavia, se l'umidità scende al di sotto di 50%, si smette di calcolare sia lo sviluppo delle spore che l'infezione.
Letteratura
- Li, Z., Dos Santos, R. F., Gao, L., Chang, P., & Wang, X. (2021). Stato attuale e prospettive future dell'antracnosi della vite causata da Elsinoe ampelina: Una malattia importante nelle regioni viticole umide. Patologia vegetale molecolare, 22(8), 899-910.
Agente patogeno
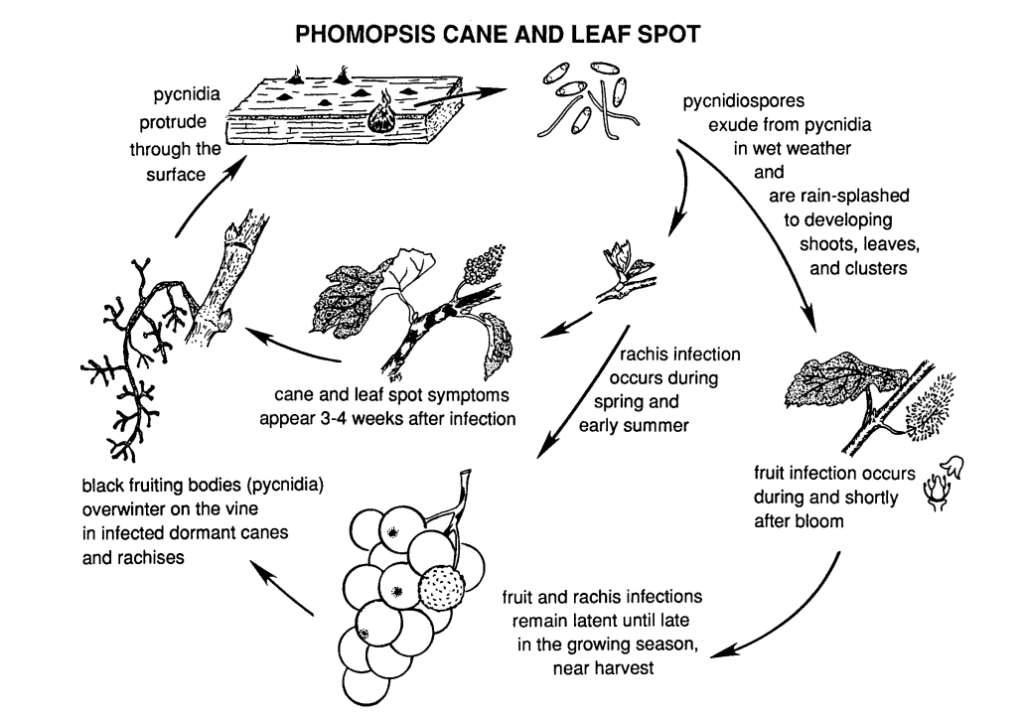
La Phomopsis delle canne e delle foglie della vite è causata da Phomopsis viticola.
Il fungo sverna nelle parti legnose della vite e torna attivo nella primavera successiva. Quando le temperature si alzano e iniziano le piogge primaverili, le spore vengono rilasciate da strutture svernanti chiamate picnidi. Quando le spore si disperdono attraverso la pioggia e il vento su nuovi tessuti, l'infezione si diffonde.
Il periodo di infezione primaria coincide tipicamente con le piogge di inizio primavera, iniziando poco dopo la rottura delle gemme, quando i germogli presentano una fase iniziale di crescita. I giovani germogli, le punte dei germogli, i grappoli di frutta e il rachide sono tutti vulnerabili all'infezione. Anche se gli apici dei germogli possono essere infettati durante tutta la stagione vegetativa, le infezioni sono più comuni dall'inizio del germogliamento alla fioritura. Nei frutti, il patogeno può rimanere inattivo fino alla maturazione dei frutti. Il periodo di incubazione dura in genere dalle 3 alle 4 settimane.
Sintomi
Sulle foglie e sui piccioli emergono piccole macchie scure con centri neri circondati da margini gialli, che alla fine si coalizzano. Le foglie basali possono deformarsi e accartocciarsi e non raggiungere le dimensioni desiderate. Quando i piccioli sono fortemente infettati, ingialliscono e cadono, portando alla caduta delle foglie. Le foglie creano un effetto ombrello che riduce ulteriori infezioni da parte delle spore.
Macchie simili compaiono sui germogli, causando la rottura degli strati epidermici nei punti di infezione. Man mano che le macchie si uniscono, le aree infette dei germogli sviluppano una consistenza tignosa. Nei casi più gravi, i germogli possono diventare stentati, rompersi o addirittura morire.
Le lesioni sul rachide lo rendono infossato e fragile. Dopo un periodo di quiescenza in estate, il fungo si riattiva all'inizio dell'autunno, provocando il marciume degli acini e dei grappoli. Sebbene le infezioni dei frutti non siano generalmente estese, sugli acini si formano macchie, spesso accompagnate da puntini neri (picnidi) sulla buccia dell'acino. In condizioni di umidità, le masse di spore gialle possono trasudare dai picnidi, causando lo scolorimento dei frutti, la loro consistenza ruvida e la mummificazione.
Il legno infetto presenta un aspetto sbiancato durante la stagione di quiescenza. Le canne e gli speroni gravemente infetti presentano decolorazioni scure miste ad aree sbiancate nella corteccia. I picinidi possono anche penetrare in superficie. Le basse temperature, insieme alla compromissione delle funzioni del floema e dello xilema, indeboliscono e uccidono le canne, gli speroni e le gemme.
Modello FieldClimate
Modello di infezione da Phomopsis in viticoltura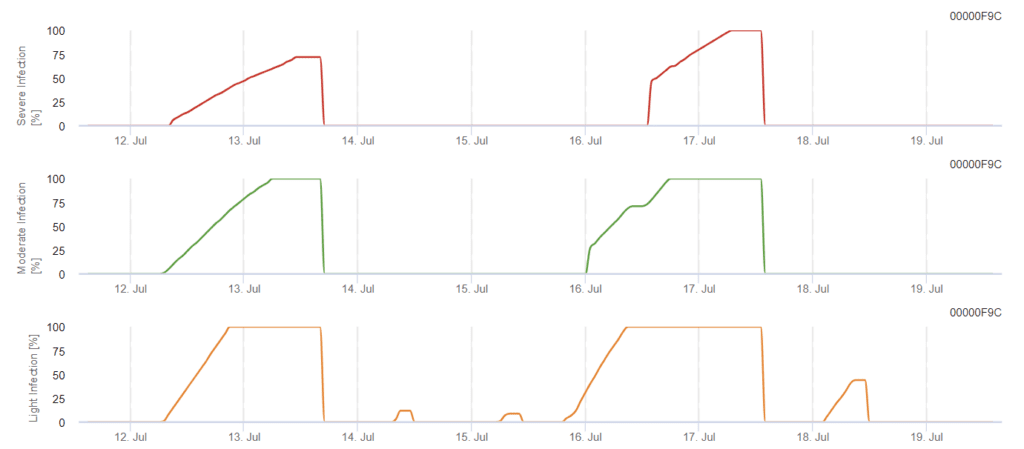
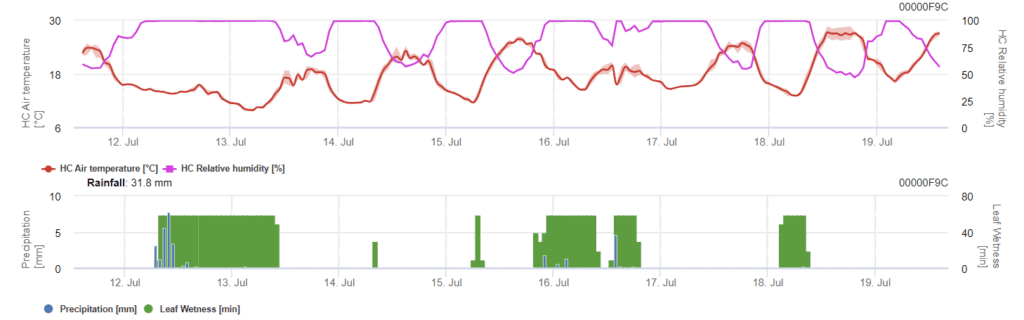
Sensori necessari:
- Temperatura dell'aria
- Umidità relativa
- Bagnatura delle foglie
- Precipitazioni
Il modello determina il rischio di infezione da Phomopsis in un intervallo di temperatura compreso tra 5 e 35°C e in condizioni di umidità (bagnatura delle foglie, elevata umidità relativa). La gravità dell'infezione dipende dalla quantità di pioggia (più di 2 mm), poiché le spore si distribuiscono più velocemente al materiale vegetale sano. Quando viene mostrata l'infezione da 100%, indica che in campo sono state misurate le condizioni ottimali per infettare i tessuti vegetali e quindi è necessario prendere in considerazione le misure di protezione delle piante.
Letteratura
- https://agriculture.vic.gov.au/biosecurity/plant-diseases/grapevine-diseases/phomopsis-cane-and-leaf-spot-of-grapevines
- Bettiga, L. J. (a cura di). (2013). Gestione dei parassiti della vite (Vol. 3343). Pubblicazioni UCANR.
- Pscheidt, J. W., & Pearson, R. C. (1991). Phomopsis cane e macchia fogliare.