

Časná plíseň brambor a rajčat
Randall C. Rowe, Sally A. Miller, Richard M. Riedel, Ohio State University Extension Service
Raná plíseň je velmi častá choroba brambor i rajčat. U brambor způsobuje skvrnitost listů a hlíz a u rajčat skvrnitost listů, hnilobu plodů a poškození stonků. Choroba se může vyskytovat v širokém spektru klimatických podmínek a může být velmi ničivá, pokud není kontrolována, a často vede k úplné defoliaci rostlin. V rozporu s názvem se zřídkakdy objevuje brzy, ale obvykle se objevuje na zralých listech.
Příznaky
Na listech obou plodin se první příznaky obvykle objevují na starších listech a skládají se z malých, nepravidelných, tmavě hnědých až černých mrtvých skvrn o velikosti od špendlíku až po půl palce v průměru. Jak se skvrny zvětšují, mohou se v důsledku nepravidelného růstu organismu v listové tkáni vytvářet soustředné kruhy. To dává poškození charakteristický vzhled "cílové skvrny" nebo "býčího oka". Kolem každé skvrny je často úzká žlutá aureola a léze jsou obvykle ohraničeny žilkami. Pokud je skvrn mnoho, mohou se sčítat, což způsobuje žloutnutí a odumírání napadených listů. Obvykle jsou nejprve napadeny nejstarší listy, které zasychají a opadávají z rostliny, jak choroba postupuje vzhůru po hlavním stonku.
Na rajčatech se infekce stonku může objevit v jakémkoli věku, což vede k malým, tmavým, mírně vpadlým plochám, které se zvětšují a vytvářejí kruhové nebo podlouhlé skvrny se světlejším středem. Na stonkových lézích se často objevují soustředné skvrny podobné těm na listech. Pokud se k založení přesazených rajčat použije napadené osivo, mohou sazenice brzy po vzejití zvlhnout. Pokud se na stoncích přesazených rostlin nebo sazenic objeví u země velké léze, mohou být rostliny opásány, což je známé jako "límcová hniloba". Takové rostliny mohou při nasazení na pole uhynout nebo, pokud jsou stonky oslabené, se mohou na začátku sezóny zlomit. Některé rostliny mohou přežít s redukovaným kořenovým systémem, pokud části stonků nad rakovinou vytvoří kořeny v místě kontaktu s půdou. Takové rostliny však obvykle plodí málo nebo vůbec. Poškození stonků je u brambor mnohem méně časté a ničivé.
Při napadení rajčat ve fázi kvetení může dojít k opadu květů, skvrnitosti stopek plodů a ztrátě mladých plodů. Na starších plodech způsobuje raná plíseň tmavé, kožovité vpadlé skvrny, obvykle v místě uchycení stopky. Tyto skvrny se mohou zvětšovat a pokrývat celou horní část plodu, často se na nich objevují soustředné znaky jako na listech. Postižená místa mohou být pokryta sametově černými výtrusy. Plody mohou být infikovány také ve fázi zelených nebo zralých plodů skrze růstové trhliny a jiné rány. Napadené plody často opadávají před dosažením zralosti.
Na hlízách brambor se časná plíseň projevuje povrchovými lézemi, které jsou o něco tmavší než přilehlá zdravá slupka. Léze jsou obvykle mírně vpadlé, kruhové nebo nepravidelné a mají různou velikost až do 3/4 palce v průměru. Mezi zdravým a nemocným pletivem je obvykle dobře ohraničený a někdy mírně vyvýšený okraj. Uvnitř tkáně je patrná hnědá až černá korkovitá suchá hniloba, obvykle ne hlubší než 1/4 až 3/8 palce. U starších lézí se mohou vytvořit hluboké trhliny. Infekce hlíz je v podmínkách Ohia vzácná.
Patogen
Raná plíseň je způsobena houbou Alternaria solani, která přežívá v infikovaných tkáních listů nebo stonků na půdě nebo v půdě. Tato houba je všeobecně přítomna na polích, kde se tyto plodiny pěstují. Může být také přenášena na semenech rajčat a v hlízách brambor. Spory se tvoří na napadených rostlinných zbytcích na povrchu půdy nebo na aktivních lézích v poměrně širokém rozmezí teplot, zejména za střídání vlhkých a suchých podmínek. Snadno se přenášejí vzdušnými proudy, větrem unášenou půdou, stříkajícím deštěm a zavlažovací vodou. K infekci náchylných pletiv listů nebo stonků dochází za teplého a vlhkého počasí s vydatnými rosami nebo deštěm. Časná plíseň se může poměrně rychle rozvinout v polovině až na konci sezóny a je silnější, pokud jsou rostliny stresovány špatnou výživou, suchem nebo jinými škůdci. K infekci hlíz brambor dochází přirozenými otvory na slupce nebo poraněním. Hlízy mohou přijít do kontaktu se sporami během sklizně a léze se mohou vyvíjet i při skladování.
TOMCAST (Jim Jasinski, koordinátor TOMCAST pro OHIO, INDIANA a MICHIGAN)
Pozadí
TOMCAST (TOMato disease foreCASTing) je počítačový model založený na polních datech, který se snaží předpovědět vývoj houbových chorob, konkrétně rané skvrnitosti, septoriové skvrnitosti listů a antraknózy rajčat. Na poli umístěné záznamníky dat zaznamenávají hodinové údaje o vlhkosti listů a teplotě. Tyto údaje byly analyzovány po dobu 24 hodin a mohou vést k tomu, že vytvoření hodnoty závažnosti onemocnění (DSV); v podstatě jde o přírůstek vývoje onemocnění. Jak se DSV hromadí, tlak chorob na plodinu stále narůstá. Pokud počet nahromaděných DSV překročí interval postřiku, doporučuje se aplikace fungicidu ke zmírnění tlaku choroby.
TOMCAST
Načasování aplikace fungicidů proti rané skvrnitosti, septoriové skvrnitosti listů a antraknóze
Systém předpovědi chorob na základě počasí nazvaný TOMCAST, který vyvinul Dr. Ron Pitblado na Ridgetown College of Agricultural Technology v kanadském Ontariu, lze použít k načasování aplikace fungicidů proti třem houbovým chorobám: rané skvrnitosti (způsobené Alternaria solani), listové skvrnitosti Septoria (způsobené Septoria lycopersici) a antraknóze plodů (způsobené Colletotrichum coccodes). Pokud se ve vašem okrese nebo v sousedních okresech vyskytuje pozdní plíseň nebo jsou podmínky pro pohyb spor do vaší oblasti, použijte k načasování aplikace fungicidů systém předpovědi pozdní plísně Simcast.
DŮLEŽITÉ UPOZORNĚNÍ:
TOMCAST není užitečný na farmách, kde se v minulosti vyskytla bakteriální onemocnění. Pokud máte běžně problémy s bakteriální skvrnitostí, skvrnitostí nebo rakovinou rajčat, neměli byste přípravek TOMCAST používat, protože doporučené intervaly postřiků nebudou dostatečné pro kontrolu bakteriálních chorob, pokud mícháte měď s aplikacemi fungicidů. Informace o omezení bakteriálních chorob naleznete zde: http://extension.psu.edu/plants/vegetable-fruit/news/2015/farming-like-you-expect-bacterial-diseases
TOMCAST používá údaje o vlhkosti listů a teplotě k výpočtu hodnot závažnosti choroby (DSV), jak je uvedeno v tabulce 1.
TOMCAST je odvozen z původního modelu F.A.S.T. (Forecasting Alternaria solani on Tomatoes), který vyvinuli doktoři Madden, Pennypacker a MacNab ? na Pennsylvania State University (PSU). Model F.A.S.T. z PSU byl dále upraven Dr. Pitbladem na Ridgetown College v Ontariu do podoby, kterou nyní známe jako model TOMCAST používaný Ohio State University Extension.
DSV jsou Hodnota závažnosti choroby (DSV) je měrnou jednotkou pro určitý stupeň rozvoje choroby (rané plísně). Jinými slovy, DSV je číselné vyjádření toho, jak rychle nebo pomalu se choroba (raná plíseň) na poli rajčat hromadí. DSV se určuje na základě dvou faktorů: vlhkosti listů a teploty během "vlhkých listů". S rostoucím počtem hodin listové vlhkosti a teplotou se DSV hromadí rychleji. Viz níže uvedený graf závažnosti choroby.
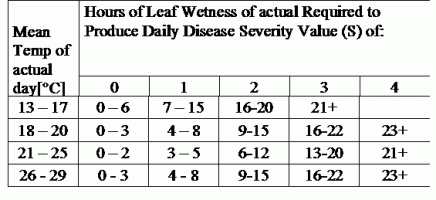
Naopak při menším počtu vlhkých hodin na listech a nižší teplotě se DSV hromadí pomalu, pokud vůbec. Když celkový počet nahromaděných DSV překročí současnou hranici, která se nazývá interval postřiku nebo práh, doporučuje se postřik fungicidem, aby se ochránily listy a plody před rozvojem choroby.

Interval postřiku (který určuje, kdy byste měli postřik provést) se může pohybovat v rozmezí 15-20 DSV. Přesnou hodnotu DSV, kterou by měl pěstitel použít, obvykle dodává zpracovatel a závisí na kvalitě plodů a konečném využití rajčat. Dodržování intervalu postřiku 15 DSV představuje konzervativní použití systému TOMCAST, což znamená, že budete postřikovat častěji než pěstitel, který používá interval postřiku 19 DSV se systémem TOMCAST. Kompromis spočívá v počtu postřiků aplikovaných během sezóny a v potenciálním rozdílu v kvalitě plodů.
POUŽÍVÁNÍ TOMCASTU
Rajčata pěstovaná v okruhu 10 mil od ohlašovací stanice by měla využívat funkci TOMCAST pro řízení chorob, která pomáhá předpovídat výskyt rané plísně, septoriózy a antraknózy. Pokud se rozhodnete vyzkoušet v této sezóně systém TOMCAST, mějte na paměti tři velmi důležité pojmy.
1) Pokud systém používáte poprvé, doporučujeme, abyste do programu zařadili pouze část své výměry, abyste zjistili, jak vyhovuje vašim standardům kvality a stylu práce.
2) Použijte TOMCAST jako vodítko, které vám pomůže lépe načasovat aplikaci fungicidů, přičemž si uvědomte, že v některých ročních obdobích můžete ve skutečnosti aplikovat více přípravku, než vyžaduje stanovený program.
3) Čím dále je pole od ohlašovacího místa, tím se zvyšuje pravděpodobnost zkreslení kumulace DSV, tj. ohlášená hodnota může být o několik DSV vyšší nebo nižší, než jaká je v místě pole. To je třeba vzít v úvahu, pokud je aplikace fungicidů pravděpodobně vzdálena několik dní. Poslouchejte hlášení DSV z okolních stanic a triangulujte s vlastní lokalitou, což je nejlepší způsob, jak zhruba odhadnout akumulaci DSV.
PRVNÍ POSTŘIK POMOCÍ TOMCASTU
V průběhu let se vedly diskuse o použití prvního postřiku při sledování přípravku TOMCAST. Pravidlo uvedené v Příručce pro produkci zeleniny z roku 1997 se soustředí na termín výsadby.
Rostliny rajčat, které vstoupí na pole před 20. květnem, by měly být poprvé postříkány, když DSV pro danou oblast překročí 25 nebo když nastane bezpečné datum 15. června. Termín fail safe se používá pouze v případě, že jste neošetřovali od 20. května, a je prostředkem k eliminaci počátečního inokula choroby. Po prvním postřiku jsou tato rajčata následně ošetřena, když je překročen zvolený interval postřiku (rozsah 15-20 DSV).
Rajčata vysazená po 20. květnu jsou ošetřena, pokud překročí zvolený interval postřiku (v rozmezí 15-20 DSV) nebo pokud nebyla ošetřena do 15. června. Proto je pro rozhodování o postřiku zásadní porovnat datum výsadby rajčat s datem zahájení hlášení DSV v dané oblasti).
Existují dvě hlavní choroby pšenice způsobené septoriózou. Jedná se o skvrnitost Septoria tritici, kterou vyvolává houba Septoria tritici (teleomorf: Mycophaerella graminicola) a Septoria nodorum skvrnitost způsobená houbou Septoria nodorum (teleomorf: Leptosphaeria nodorum). Obě choroby způsobují vážné ztráty na výnosech, které se údajně pohybují od 31 do 53 % (Eyal, 1981; Babadoost a Herbert, 1984; Polley a Thomas, 1991). Celosvětově je napadeno více než 50 milionů ha pšenice, která roste především v oblastech s vysokými srážkami. Během posledních 25 let se počet těchto chorob zvyšuje a v některých oblastech se staly hlavním limitujícím faktorem produkce pšenice. Při silných epidemiích jsou zrna náchylných odrůd pšenice scvrklá a nejsou vhodná k mletí. Epidemie skvrnitosti pšenice Septoria tritici a Septoria nodorum jsou spojeny s příznivými povětrnostními podmínkami (časté deště a mírné teploty), specifickými pěstitelskými postupy, dostupností inokula a přítomností náchylných odrůd pšenice (Eyal et al., 1987).
Septoria spp. Biologie
Za Erickem De Wolfem, Septoria Tritici Blotch, Kansas State University, duben 2008 Septoria tritici známá jako skvrnitá skvrnitost listů, je způsobena houbou Septoria tritici. Je rozšířena ve všech oblastech pěstování pšenice na světě a v mnoha regionech představuje vážný problém. Největší škody způsobuje skvrnitost Septoria tritici, když choroba napadá horní listy a hlávky náchylných odrůd v pozdní fázi sezóny.
Příznaky
Septoria tritici příznaky skvrnitosti se poprvé objevují na podzim. Prvními příznaky jsou malé žluté skvrny na listech. Tyto léze se stárnutím často zbarvují do světle hnědé barvy a na řapících lze pozorovat plodnice houby usazené v lézích. Léze mají nepravidelný tvar a jsou od eliptických až po dlouhé a úzké (obrázek 1). Léze obsahují malé kulaté černé skvrny, které jsou plodnicemi houby. Černé plodnice vypadají jako zrnka černého pepře a obvykle je lze spatřit bez použití lupy. Choroba začíná na spodních listech a postupně se šíří na praporcovité listy. K napadení jsou náchylné i listové pochvy. Ve vlhkých letech se může houba skvrnitost listů přesunout na hlávky a způsobit hnědé léze na lodyhách a řapících, známé jako skvrnitost lodyh. Tyto léze se stárnutím často zbarvují do světle hnědé barvy a v lézích na řapících jsou často vidět plodnice houby.
Fáze skvrnitosti lusků může způsobit značné ztráty na výnosech, ale vztah mezi závažností choroby a ztrátami na výnosech není dobře znám. Septoria tritici skvrnitost může být zaměněna s jinými chorobami listů, které mají velmi podobné příznaky: opálení a skvrnitost listů. Stagonspora nodorum například skvrny. Je běžné, že rostliny jsou napadeny více než jednou z těchto listových chorob, a pro přesnou diagnózu, které choroby převažují, může být nutné laboratorní vyšetření. K rozlišení příčiny skvrnitosti listů je téměř vždy nutné laboratorní vyšetření. Znalost druhu není pro rozhodnutí o postřiku důležitá, protože všechny tři choroby reagují na fungicidy podobně. Znalost toho, které choroby jsou nejrozšířenější, je však důležitou součástí výběru odrůd, protože odolnost vůči chorobám řídí různé geny.
Nejspolehlivějším způsobem, jak odlišit skvrnitost Septoria tritici od ostatních chorob, je přítomnost černých plodnic. Houba, která způsobuje opálenou skvrnitost, tento typ rozmnožovací struktury nevytváří. Houba způsobující skvrnitost Stagonospora nodorum však za vlhkých podmínek vytváří světle hnědé plodnice. Kromě barevného rozdílu jsou tyto struktury také menší než struktury vytvářené houbou Septoria tritici.
Životní cyklus
Septoria tritici přežívá přes léto na zbytcích předchozí úrody pšenice a na podzim iniciuje infekci. Existují určité důkazy o tom, že houba je schopna přežívat ve spojení s jinými hostiteli trav a osivem pšenice. Tyto zdroje houby jsou pravděpodobně nejdůležitější v době, kdy zbytky pšenice chybí. Bez ohledu na střídání plodin nebo postupy hospodaření s rezidui je obvykle dostatek inokula k iniciaci podzimních infekcí. Septoria tritici skvrnitosti svědčí chladné a vlhké počasí. Optimální teplota je 16 až 21 °C, avšak k infekci může dojít i v zimních měsících při teplotách do 5 °C. Infekce vyžaduje alespoň 6 hodin vlhka v listech a pro maximální infekci je zapotřebí až 48 hodin vlhka. Po infekci trvá houbě 21 až 28 dní, než vytvoří charakteristické černé plodnice a vyprodukuje novou generaci spor. Výtrusy vytvořené v těchto plodnicích jsou vylučovány v lepkavé hmotě a vyžadují déšť, aby se rozstříkly na horní listy a hlávky.
Infekce způsobená Septoria tritici
Pycnidiospory S. tritici klíčí ve volné vodě z obou konců výtrusů nebo z interkalárních buněk (Weber, 1922). Výtrusy začínají klíčit až asi 12 hodin po kontaktu s listem. Zárodečné trubice rostou náhodně po povrchu listu. Weber (1922) pozoroval pouze přímý průnik mezi epidermálními buňkami, ale jiní dospěli k závěru, že primárním způsobem průniku do hostitele je průnik přes otevřená i uzavřená stomata (Benedict, 1971; Cohen a Eyal, 1993; Hilu a Bever, 1957). Kema et al. (1996) pozorovali pouze průnik přes stomy. Hyfy prorůstající stomaty se zužují na průměr asi 1 μm a po dosažení substomatální dutiny se rozšiřují.
Hyfy rostou rovnoběžně s povrchem listu pod epidermálními buňkami, pak přes mezofyl k buňkám spodní epidermis, ale ne do epidermis. Netvoří se haustoria a růst hyf je omezen sklerenchymatickými buňkami kolem cévních svazků, s výjimkou případů, kdy jsou hyfy velmi husté. Cévní svazky nejsou napadeny. Hyfy prorůstají mezibuněčně podél buněčných stěn mezofylem a větví se na přepážce nebo uprostřed buňky. Přibližně 9 dní se neobjevují žádné makroskopické příznaky s výjimkou občasné mrtvé buňky, ale po 11 dnech buňky mezofylu rychle odumírají. Pyknidy se vyvíjejí v substomatálních komůrkách. Hyfy zřídka prorůstají do buněk hostitele (Hilu a Bever, 1957; Kema et al, 1996; Weber, 1922).
K úspěšné infekci dochází až po nejméně 20 hodinách vysoké vlhkosti. Pouze několik hnědých skvrn se vyvinulo, pokud listy zůstaly vlhké 5 až 10 hodin po uložení spor (Holmes a Colhoun, 1974) nebo až 24 hodin (Kema et al., 1996). Vztahy mezi hostitelem a parazitem jsou stejné na rezistentní i náchylné pšenici. Klíčení spor na povrchu listů je stejné bez ohledu na citlivost. Počet úspěšných průniků je přibližně stejný, ale růst hyf je rychlejší u náchylných odrůd, což vede k většímu počtu lézí. Hyfy přesahují 44 Session 2 - B.M. Cunfer nekrotickou oblast u všech odrůd. V patogenezi může hrát roli toxin (Cohen a Eyal, 1993; Hilu a Bever, 1957). Naproti tomu na rezistentní linii byla kolonizace značně omezena (Kema et al., 1996).
Stagonospora (Septoria) a Septoria Pathogens of Cereals: Proces infekce
B.M. Cunfer, Department of Plant Pathology, University of Georgia, Griffin, GA
Proces infekce byla nejintenzivněji studována pro Stagonospora (Septoria) nodorum a Septoria tritici. Jedna hloubková studie o Septoria passerinii je k dispozici. Téměř všechny uváděné informace se týkají infekce pyknidiosporami. Proces infekce u jiných forem spor je však dosti podobný. Uvedené informace se většinou týkají infekce listů za optimálních podmínek. Některé studie byly provedeny s neporušenými semenáčky, zatímco jiné s oddělenými listy. Infekce koleoptile a semenáčků pšenice pomocí S. nodorum podrobně popsal Baker (1971) a recenzoval Cunfer (1983). Ačkoli nebylo provedeno žádné přesné srovnání, zdá se, že proces infekce má mnoho podobností v každém systému hostitel-parazit a je typický pro mnoho nekrotrofních patogenů. Informace o faktorech ovlivňujících vývoj symptomů a projevy choroby jsou vyloučeny, ale byly přezkoumány jinými autory (Eyal et al., 1987; King et al., 1983; Shipton et al., 1971). Je zahrnut přehled faktorů ovlivňujících životnost spor na povrchu listů.
Úloha Cirrusu a přežití spor na povrchu listů Nejpodrobnější informace o funkci cirru, který obaluje pyknidiospory vylučované z pyknidia, se týkají S. nodorum. Cirrus je gel složený z bílkovinných a sacharidových sloučenin. Jeho složení a funkce jsou podobné jako u jiných hub z čeledi Sphaeropsidales (Fournet, 1969; Fournet et al., 1970; Griffiths a Peverett, 1980). Primárními úlohami cirrových složek jsou ochrana pyknidiospor před vysycháním a prevence předčasného klíčení.
Cirrus chrání pyknidiospory, takže některé z nich zůstávají životaschopné nejméně 28 dní (Fournet, 1969). Když byl cirrus zředěn vodou, pokud byla koncentrace roztoku cirrusu >20%, vyklíčilo méně než 10% pyknidiospor. Při nižší koncentraci poskytují složky živiny, které stimulují klíčení spor a prodlužování zárodečných trubic. Délka zárodečných trubic se zvyšovala až do koncentrace 15% cirru, poté při vyšších koncentracích mírně klesala (Harrower, 1976). Brennan et al. (1986) zaznamenali větší klíčivost ve zředěné cirrusové tekutině. Složky cirru snižovaly klíčivost při relativní vlhkosti 10-60%. Po rozptýlení spor jsou stimulační účinky cirrusové tekutiny pravděpodobně zanedbatelné (Griffiths a Peverett, 1980).
Při relativní vlhkosti 35-45% jsou spory S. tritici v cirri zůstaly životaschopné nejméně 60 dní (Gough a Lee, 1985). Složky cirry mohou působit jako inhibitor klíčení spor nebo může klíčení bránit vysoký osmotický potenciál cirry. Pycnidiospory S. nodorum nepřežily 24 hodin při relativní vlhkosti vzduchu nad 80% při 20 C. Spory přežívaly dva týdny nebo déle při relativní vlhkosti <10% (Griffiths a Peverett, 1980). Když se cirrusová tekutina S. nodorum byla zředěna vodou, přibližně dvě třetiny pyknidiospor ztratily životaschopnost během 8 hodin a po 30 hodinách na denním světle vyklíčilo pouze 5%. Při skladování spor ve tmě zůstalo 40% životaschopných i po 30 hodinách (Brennan et al., 1986).
Suché konidie S. nodorum, ve stínu i na přímém slunci, přežily venku nejméně 56 hodin (Fernandes a Hendrix, 1986a). Klíčení pyknidiospor S. nodorum bylo inhibováno kontinuálním UV-B zářením (280-320 nm), zatímco klíčení pyknidiospor S. nodorum bylo inhibováno kontinuálním UV-B zářením (280-320 nm). S. tritici nebyla. Prodlužování zárodečných trubic pod nepřetržitým UV-B zářením bylo u obou hub ve srovnání s tmou inhibováno (Rasanayagam et al., 1995).
Infekce způsobená Septoria nodorum
Proces pronikání do hostitele a vývoj S. nodorum uvnitř listu podrobně zkoumalo několik badatelů (Baker a Smith, 1978, Bird a Ride 1981, Karjalainen a Lounatmaa, 1986; Keon a Hargreaves, 1984; Straley, 1979; Weber, 1922). Pyknidiospory mají tendenci ulpívat v prohlubních mezi dvěma epidermálními buňkami a mnoho pokusů o průnik do listů začíná právě tam. Spory klíčí na povrchu listů v reakci na volnou vlhkost (Fernandes a Hendrix, 1986b). Začínají klíčit 2-3 hodiny po uložení a po 8 hodinách může klíčivost dosáhnout 90%. Pronikání do listů začíná asi 10 hodin po uložení spor (Bird a Ride, 1981; Brönnimann a kol., 1972; Holmes a Colhoun, 1974).
Na počátku klíčení je zárodečná trubice obklopena amorfním materiálem, který se přichytí k listu. Zárodečné trubice vyrůstající z obou konců výtrusu a z interkalárních buněk mají tendenci růst podél prohlubní mezi buňkami a jsou často orientovány podél dlouhé osy listu (O'Reilly a Downes, 1986). Hyfy ze spor, které nejsou v prohlubních, rostou náhodně s občasným větvením (Straley, 1979). Vytváří se apresorium s infekčním kolíkem, který proniká kutikulou a periklinálními stěnami epidermálních buněk přímo do buněčného lumen, což vede k rychlé buněčné smrti.
Mnoho průniků je nejprve subkutikulárních nebo dochází k bočnímu růstu hyfy v buněčné stěně před růstem do cytoplazmy (Bird a Ride, 1981; O'Reilly a Downes, 1986). Dochází také k průniku skrz otevřené i uzavřené žaludky, který může být rychlejší než přímý průnik (Harrower, 1976; Jenkins, 1978; O'Reilly a Downes, 1986; Straley, 1979). Zárodečné trubice se rozvětvují u žaludů a spojů epidermálních buněk. K průniku zárodečné trubice do stomatu může dojít bez vytvoření apresoria. Někdy dochází k průniku přes trichomy (Straley, 1979). Většina pokusů o průnik je zřejmě neúspěšná, v buňkách v místě pokusu o průnik se vytvářejí husté papily (Karjalainen a Lounatmaa, 1986; Bird a Ride, 1981).
Po proniknutí epidermální buňky rychle odumírají a lignifikují a hyfy prorůstají do mezofylu. Buňky mezofylu se deformují a lignifikovaný materiál se ukládá vně některých buněk, které pak zkolabují. K lignifikaci dochází dříve, než hyfy dosáhnou buňky. Tento proces je u všech odrůd stejný, ale u odolných odrůd se vyvíjí pomaleji. Hyfy prorůstají mezibuněčně mezi epidermálními buňkami a poté do mezofylu. Po proniknutí do mezofylu začíná za 6-9 dní degradace chloroplastů (Karjalainen a Lounatmaa, 1986).
Rychlost fotosyntézy však začíná klesat během jednoho dne po infekci a dříve, než se projeví příznaky (Krupinsky et al, 1973). Sklerenchymatické pletivo kolem cévních svazků zabraňuje infekci cévního pletiva. Cévní svazky blokují šíření hyf mezofylem s výjimkou případů, kdy je sklerenchymatické pletivo mladé a není plně vytvořeno (Baker a Smith, 1978).
Stagonospora nodorum uvolňuje in vitro a během infekce listů pšenice širokou škálu enzymů degradujících buněčnou stěnu, včetně amylázy, pektinmetylesterázy, polygalakturonázy, xylanázy a celulázy (Baker, 1969; Lehtinen, 1993; Magro, 1984). Informace týkající se degradace buněčných stěn enzymy se shodují s histologickými pozorováními. tyto enzymy mohou působit ve spojení s toxiny. Citlivost na enzymy může souviset s odolností a rychlostí kolonizace houbami (Magro, 1984). Stejně jako mnoho nekrotrofů produkují patogeny Septoria a Stagonospora fytotoxické sloučeniny in vitro. Zničení a odumření buněk před prorůstáním hyf do mezofylového pletiva (Bird a Ride, 1981) odpovídá produkci toxinů. Definitivní úloha toxinů v procesu infekce a jejich vztah k odolnosti hostitele však nebyla stanovena (Bethenod et al, 1982; Bousquet et al, 1980; Essad a Bousquet, 1981; King et al, 1983). Rozdíly v hostitelském spektru mezi kmeny adaptovanými na pšenici a ječmen S. nodorum může souviset s produkcí toxinů (Bousquet a Kollmann, 1998). Iniciace klíčení spor a procento vyklíčených spor nejsou ovlivněny citlivostí hostitele (Bird a Ride, 1981; Morgan 1974; Straley, 1979; Straley a Scharen, 1979; Baker a Smith, 1978).
Bird a Ride (1981) uvádějí, že prodlužování zárodečných trubic na povrchu listů bylo pomalejší u rezistentních než u náchylných odrůd. Tento mechanismus, který se projevil nejméně 48 hodin po uložení spor, ukazuje na odolnost vůči prodlužování zárodečných trubic před průnikem. U rezistentních odrůd bylo méně úspěšných průniků a průnik probíhal na rezistentních odrůdách pomaleji (Baker a Smith, 1978; Bird a Ride, 1981). Bylo navrženo, že lignifikace omezuje infekci u rezistentních i náchylných kultivarů, ale další faktory zpomalují vývoj houby u rezistentních linií. U náchylných linií mohou rychleji rostoucí hyfy unikat lignifikaci hostitelských buněk. čtyři dny po inokulaci ječmene izolátem biotypu pšenice z. S. nodorum, hyfy prorůstaly kutikulou a někdy i vnějšími celulózovými vrstvami buněčných stěn epidermis. Pod pronikajícími hyfami byly uloženy silné papily a buňky nepronikaly (Keon a Hargreaves, 1984).
Infekce způsobená Septoria passerinii: Green a Dickson (1957) podrobně popisují proces infekce u bakterií S. passerinii na ječmeni. Proces infekce je podobný S. tritici. Stejně jako u S. tritici je doba potřebná k proniknutí do listů podstatně delší než u S. tritici. S. nodorum. Zárodečné trubice se větví a rostou po povrchu listů náhodně, ale někdy i podél prohlubní mezi epidermálními buňkami. Do listů pronikají téměř výhradně přes průduchy. Klíční hyfy nabobtnají, a pokud je průnik neúspěšný, hyfy se dále prodlužují. K pronikání nedochází 48 hodin po uložení spor. Po 72 hodinách se zárodečné trubice zahušťují přes stomata, prorůstají mezi ochrannými buňkami a na urfaces akcesorních buněk a do substomatálních dutin. Přímý průnik mezi epidermální buňky je pozorován jen výjimečně.
Klíčivost spor a pronikání do hostitele jsou stejné u rezistentních i náchylných odrůd. Na rezistentních odrůdách je mnohem menší rozšíření hyf uvnitř listů a na mnoha, ale ne všech buněčných stěnách jsou pozorovány papily. Hyfy prorůstají pod epidermis z jednoho stomu do druhého, ale nepronikají mezi epidermální buňky. Mezofyl je kolonizován, ale netvoří se haustoria. Po odumření buněk mezofylu se epidermální buňky rozpadají. Vývoj mycelia v listu je řídký a obvykle blokovaný cévními svazky. U mladších listů, pokud je cévní pochva méně vyvinutá, procházejí hyfy mezi svazkem a epidermis. Pyknidy se tvoří v substomatálních dutinách, většinou na horním povrchu listu (Green a Dickson, 1957).
Faktory ovlivňující životnost spor na povrchu listů Z patogenů Stagonospora a Septoria na obilovinách byly definitivní informace o infekčním procesu podány pouze pro tyto patogeny. S. nodorum, S. triticia S. passerinii. Stejně jako mnoho jiných nekrotrofních patogenů ani tato skupina patogenů nevyvolává hypersenzitivní reakci. Významný rozdíl v infekčním procesu mezi patogeny Septoria a Stagonospora spočívá v tom, že klíčení a pronikání spor probíhá mnohem rychleji u S. nodorum než pro S. tritici a S. passerinii. To má významný vliv na epidemiologii onemocnění.
Patogeny rodu Septoria pronikají do rostliny především skrze žaludky, kdežto S. nodorum proniká přímo i přes žaludky. S. nodorum rychle proniká do epidermálních buněk a zabíjí je, ale S. tritici a S. S. passerinii nezabíjejí epidermální buňky, dokud se hyfy nerozvětví v mezofylu listů a nezačne rychlá nekróza. Histologické studie růstu hub po proniknutí do hostitele odpovídají údajům získaným z epidemiologických studií rezistence hostitele. Rezistence zpomaluje rychlost kolonizace hostitele, ale nemá výrazný vliv na proces vývoje lézí.
Mechanismy, které řídí reakci hostitele, ať už se týkají enzymů a toxinů nebo jiných metabolitů uvolňovaných patogeny během infekce, jsou stále nejasné. O infekci askosporami je málo informací. Proces infekce je pravděpodobně velmi podobný procesu infekce pyknidiosporami. Askospory Phaeosphaeria nodorum klíčí v širokém rozmezí teplot a jejich zárodečné trubice pronikají přímo do listu. Podle Rapillyho et al. (1973) však askospory na rozdíl od pyknidiospor neklíčí ve volné vodě.
Model infekce Septoria spp.
Septoriové infekce jsou možné při nízkých teplotách, zatímco teploty pod 7 °C nemusí vést k infekci do dvou dnů. Optimální teploty pro onemocnění je dosaženo v oblasti 16 až 21 °C. Infekce jsou možné v období vysoké relativní vlhkosti vzduchu nebo vlhkosti listů trvajícím 14 hodin nebo déle. Pro splnění těchto podmínek jsme se rozhodli rozdělit na modely pro slabé, středně silné a silné infekce. Slabé infekce lze uvést, pokud je možné, aby patogen infikoval tkáň hostitele. To znamená, že ke slabým infekcím může dojít, pokud jsou teploty v minimech a doba vlhkosti listů je kritické délky. Středně silná infekce proběhne za podmínek, kdy většina infekčních pokusů vede k přiměřeným výsledkům, a silná infekce proběhne za podmínek, kdy má patogen optimální podmínky pro infekci.

Infekce začíná po dešti 0,5 mm. Rozhodli jsme se nepoužít model pro tvorbu pyknid. Předpokládáme, že podmínkou potřebnou pro tvorbu pyknid je období s relativní vlhkostí vyšší než 85%. Doba života pyknid se předpokládá 24 hodin. Ve všech klimatických podmínkách, kde má Septoria tritici šanci se infikovat, najdeme 2 hodiny splňující tuto podmínku téměř každý den kolem východu slunce.
Hodnocení závažnosti infekce: Abychom mohli posoudit infekční tlak Septoria tritici mezi fází 10 (první listový žlábek) a fází 32 (druhý uzel alespoň 2 cm nad uzlem 1) a mezi 32 a 51 (začátek hlávkování), musíme posoudit závažnost infekce na základě klimatických podmínek. Toto hodnocení se provádí ve stupnici od 1 do 5. Závažnost 1 je dána, pokud je splněna podmínka pro slabou infekci a napršelo méně než 5 mm, jinak bude odpovídající stupeň závažnosti 2. Závažnost 3 je uvedena, pokud je splněna podmínka slabé infekce a napršelo méně než 5 mm. Pokud při středně těžké infekci napršelo více než 5 mm nebo při těžké infekci méně než 5 mm, je uvedena závažnost 4.
Silná infekce s více než 5 mm srážek odpovídá hodnotě závažnosti 5.
Septoria tritici hodnocení tlaku nemoci: Podnebí je pouze jedním z faktorů, které rozhodují o tlaku chorob na poli. Dalšími dvěma faktory jsou historie pole a náchylnost pěstované odrůdy. Pokud můžeme shrnout hodnoty intenzity choroby od fáze 10 do fáze 32 do hodnoty 4, můžeme očekávat slabý tlak choroby způsobený klimatem. Pokud tato hodnota dosáhne 6, můžeme očekávat mírný tlak choroby a pokud dosáhne 10, můžeme očekávat silný tlak choroby ze strany klimatu. Znalost náchylnosti odrůdy a historie pole nás v této situaci povede k tomu, zda postřik provádět, nebo neprovádět při slabém nebo středním tlaku choroby. Mít kumulativní hodnotu 10 může vést k postřiku ve fázi 32 tak jako tak. Rozhodnutí o postřiku v pozdější fázi je více závislé na jarním klimatu. Pokud jsme schopni kumulovat hodnoty intenzity od fáze 10 na hodnotu 6, můžeme očekávat slabý tlak choroby. Pokud tato hodnota dosáhne 10, můžeme očekávat mírný tlak choroby, a pokud tato hodnota dosáhne 15, můžeme očekávat silný tlak choroby v důsledku klimatické situace.
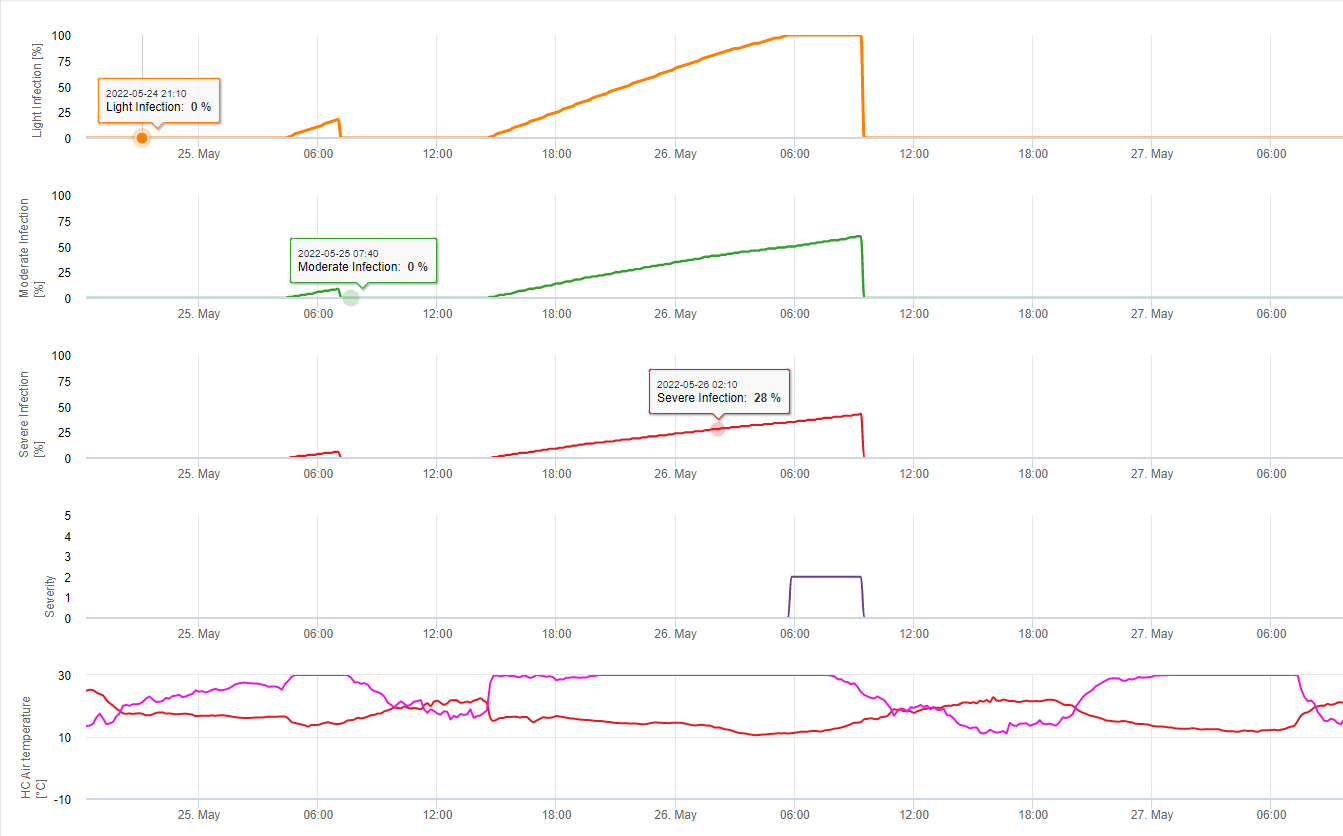
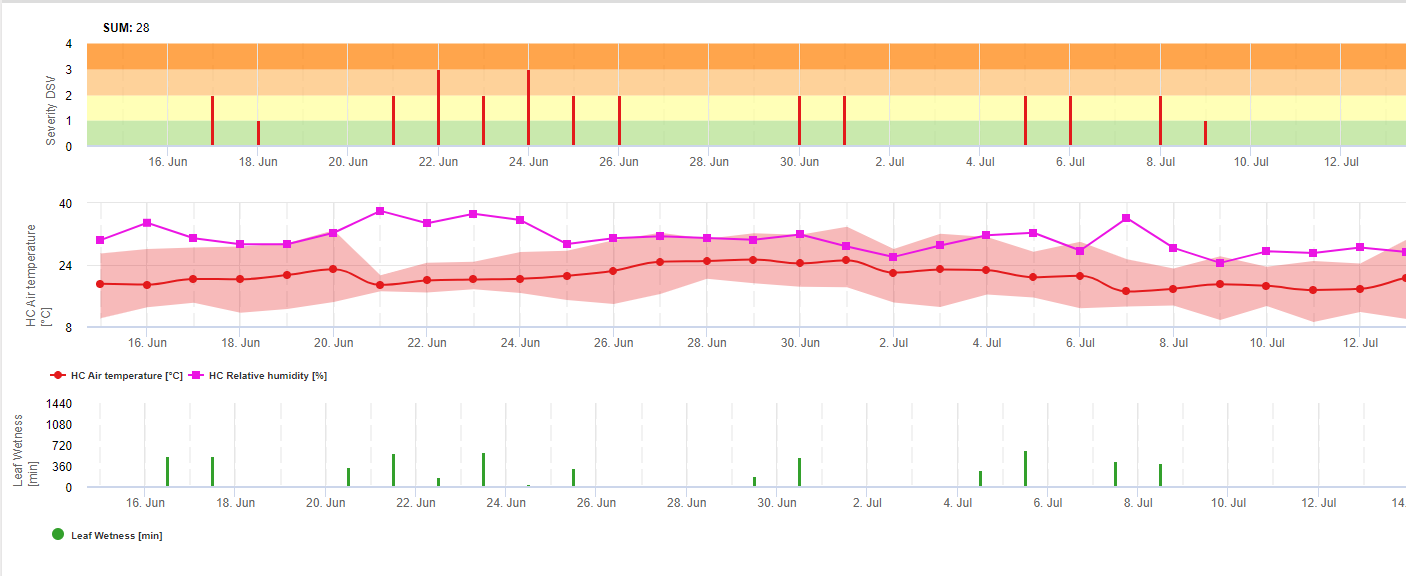
V programu FieldClimate ukazujeme Septoria tritici Závažnost společně se třemi různými závažnostmi infekce v jednom grafu (viz výše). Vzhledem k dešťovým srážkám a dlouhým obdobím vlhka listů jsou podmínky pro silnou infekci způsobenou S. tritici 14. a 16. května bylo plné pole. Úrovně závažnosti dosahují 14. května nejvyšší hodnoty 5, což znamená, že nyní hrozí vysoké riziko infekce.
Biologie infekce Stagonospora nodorum se do jisté míry liší od biologie S. tritici, ale tento rozdíl není dostatečně velký na to, aby byl vytvořen samostatný model. Proto navrhujeme použít tento model pro celý komplex stagnosporóz a septorióz v obilovinách včetně S. passerinii. S. tritici a S. passerinii potřebují delší období vlhkosti listů než S. nodorum. V oblastech s vysokým tlakem S. nodorum infekce klasifikované jako slabé s hodnotou závažnosti 2 by měly být léčeny závažněji než v jiných oblastech.
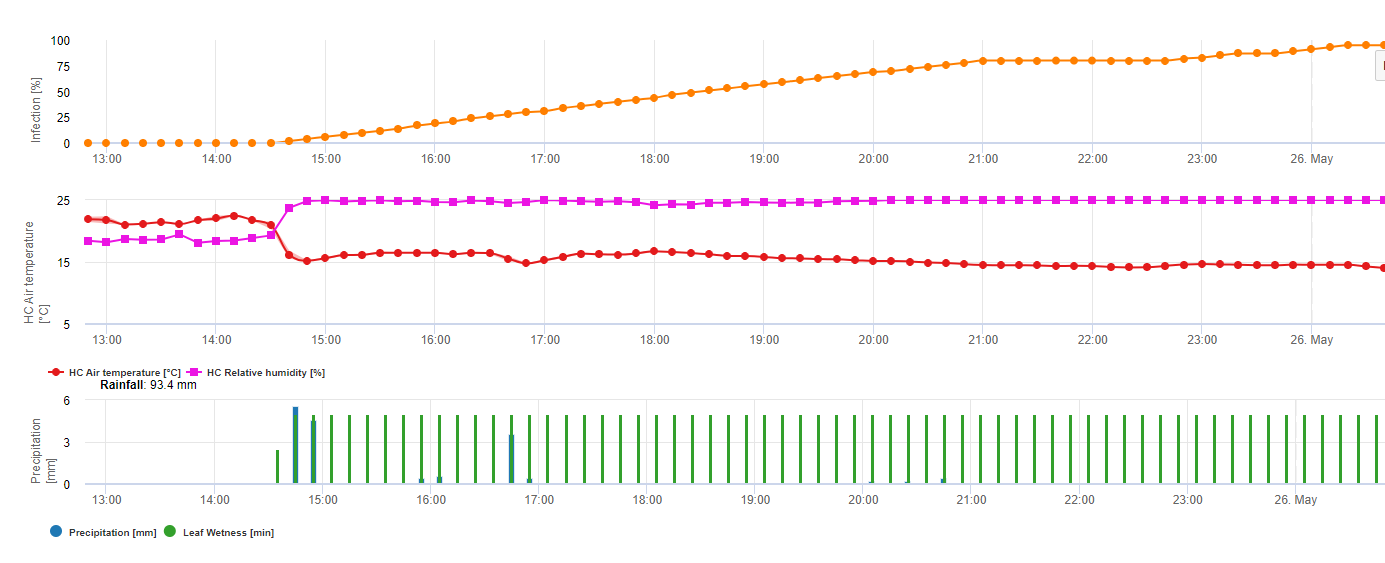
Pro Septoria nodorum rizikový model je uveden v FieldClimate (viz výše). Vysoké riziko bylo určeno na zasedání 17. června a 7. července (100%). V závislosti na stádiu náchylnosti rostlin k infekci je třeba vzít v úvahu opatření na ochranu rostlin, pokud riziko dosáhne 80% (viz také předpověď počasí, ochranná ochrana rostlin). Pokud riziko dosáhlo 100% a byla již zjištěna infekce, je třeba provést systémová opatření na ochranu rostlin (kurativní aplikace).
Několik druhů rostlinných patogenních hub z rodu Colletotrichum způsobují antraknózu paprik a mnoha dalších druhů zeleniny a ovoce. Až do konce 90. let 20. století byla antraknóza paprik a rajčat spojována pouze se zralými nebo dozrávajícími plody. Od té doby se prosadila agresivnější forma choroby. Tato forma napadá papriky v kterékoli fázi vývoje plodů a může ohrozit rentabilitu úrody paprik v oblastech, kde se rozšířila. Tato choroba může napadnout také rajčata, jahody a případně i další ovocné a zeleninové plodiny.
Příznaky
Na nezralých plodech jakékoli velikosti se objevují kruhové nebo hranaté vpadlé léze. Na jednotlivých plodech se často tvoří více lézí. Při silném výskytu choroby se mohou léze spojovat. Na povrchu lézí se často v soustředných kruzích vytvářejí růžové až oranžové masy spor houby. U starších lézí lze pozorovat černé struktury zvané acervuli. Při pohledu ručním objektivem vypadají jako malé černé tečky, pod mikroskopem jako chomáčky drobných černých chloupků. Patogen rychle a hojně vytváří spory a může se rychle rozšířit po celé úrodě papriky, což vede ke ztrátě výnosu až 100%. Poškození se mohou objevit také na stoncích a listech jako nepravidelně tvarované hnědé skvrny s tmavě hnědými okraji.
Patogen
Tuto formu antraknózy papriky způsobuje houba Colletotrichum acutatum. Patogen přežívá na rostlinných zbytcích z infikovaných plodin a na jiných náchylných druzích rostlin. Při absenci napadených rostlinných zbytků není houba dlouhodobě přenosná v půdě. Houba se může do plodiny dostat také na napadeném osivu. Během teplá a vlhká období, spory jsou rozprašovány dešťovou nebo zavlažovací vodou. z nemocných plodů na zdravé. Nemocné plody jsou zdrojem inokula, které umožňuje šíření choroby z rostliny na rostlinu na poli. Houba přežívá v semenech a na nich. Antraknóza se na pole dostává s infikovanými přesazenými rostlinami nebo může přežívat mezi sezónami v rostlinných zbytcích nebo na plevelných hostitelích. Alternativními hostiteli jsou plevele a další rostliny z čeledi Solanaceae (rajče, brambor, lilek), ačkoli infekce těchto hostitelů jsou na Floridě velmi vzácné. Plody jsou infikovány, když jsou spory houby nebo napadené zbytky deštěm rozprašovány na rostliny papriky. V infikovaném pletivu se vytvářejí nové spory, které se pak šíří na další plody. Pracovníci mohou spory přenášet také pomocí zařízení nebo při manipulaci s infikovanými rostlinami. K infekci obvykle dochází za teplého a vlhkého počasí. Teploty kolem Optimální teplota je 80° F (27° C). pro rozvoj choroby, ačkoli k infekci dochází jak při vyšších, tak při nižších teplotách. K velkým ztrátám dochází za deštivého počasí, protože výtrusy jsou vyplavovány nebo rozprašovány na další plody, což vede k většímu počtu infekcí. Choroba se s větší pravděpodobností vyvíjí na zralých plodech, které jsou na rostlině přítomny delší dobu, ačkoli se může vyskytnout jak na nezralých, tak na zralých plodech. Antraknóza může napadat při teplotách od 15 °C do 30 °C. Ale a dlouhá listová vlhkost je zapotřebí k naplnění potřeb infekce. Při optimální teplotě z 20 °C na 25 °C ještě 12 hodin po navlhčení listů jsou potřebné. Vyšší nebo nižší teplota vyžaduje ještě delší období zvlhčování listů (pro výpočet není nutná lineární funkce nebo matice). FieldClimate počítá možné infekční události na základě vlhkosti listů a teplot během této události.
Biologie Plasmopara viticola (plíseň puchýřkatá)
P. viticola je obligátní parazit. To znamená, že k růstu potřebuje zelené, čerstvé orgány révy. Během bezvegetačního období přetrvává a vytváří plodnice, tzv. oospory. Oospory oomycetů mohou v půdě přežívat velmi dlouho. Proto se s puchýřnatkou můžeme setkat i na místech, kde se infekce nevyskytují v každém roce. Na jaře, když je svrchní část půdy dostatečně vlhká a teplá, vytvoří oospory tzv. makrosporangia, která mohou uvolnit až 200 zoospor do volné vody. Zoospory se větrem v kapičkách vody přesouvají na listy a do shluků. Mají však dva bičíky a pohybují se ve vodním filmu na spodní straně listů nebo trsů a mladých bobulí, aby našly stoma a pronikly do rostlinných pletiv. Vstoupí do stomu a vyklíčí v něm, kam během necelé hodiny přenesou veškerou svou plazmu. V mikroskopických studiích bylo nalezení stomu, encystace a klíčení do stomu ukončeno během 90 minut.
P. viticola roste v mezibuněčném prostoru a živí se haustoriemi pronikajícími do epidermálních a parenchymových buněk. V závislosti na teplotě a relativní vlhkosti se vyvíjí dostatečný mezibuněčný růst s dostatečným množstvím haustorií, aby se vytvořilo substomatózní tělo, které vyplňuje celý substomatózní prostor a které vyzdvihuje epidermální tkáň z parenchymové tkáně. To vede k viditelnému příznaku olejové skvrny.
Oomycety jsou sporulující za nepřítomnosti světla, když relativní vlhkost vzduchu je velmi vysoká. Na adrese P. viticola nedochází ke sporulaci, pokud je teplota nižší než 12 °C a relativní vlhkost nižší než 95%. Sporangiafory jsou tvořeny vezikulami substomatu a vycházejí ze stomatu. Čerstvě vytvořená sporangia jsou lepkavá a lze je ze sporangiaforů odstranit pouze vodou. Při poklesu relativní vlhkosti vzduchu se sporangia stávají pokusnými a mohou být odstraněna i větrem.
Sporangia uvolní do volné vody až 20 zoospor. Tyto zoospory musí být roznášeny větrem ve vodních kapkách, aby se dostaly na čerstvé listy, nebo mohou být sporangia roznášena deštěm či větrem samotným. Proces infekce při primární a sekundární infekci je stejný.
Vzhledem k velkému významu pohlavního stadia pro hibernaci patogenu můžeme předpokládat, že ve všech vinicích, kde se vyskytuje plíseň révová, se hodí k páření. K tvorbě zoospor dochází na starších listech během léta a začátkem podzimu.
Primární infekce podrobně
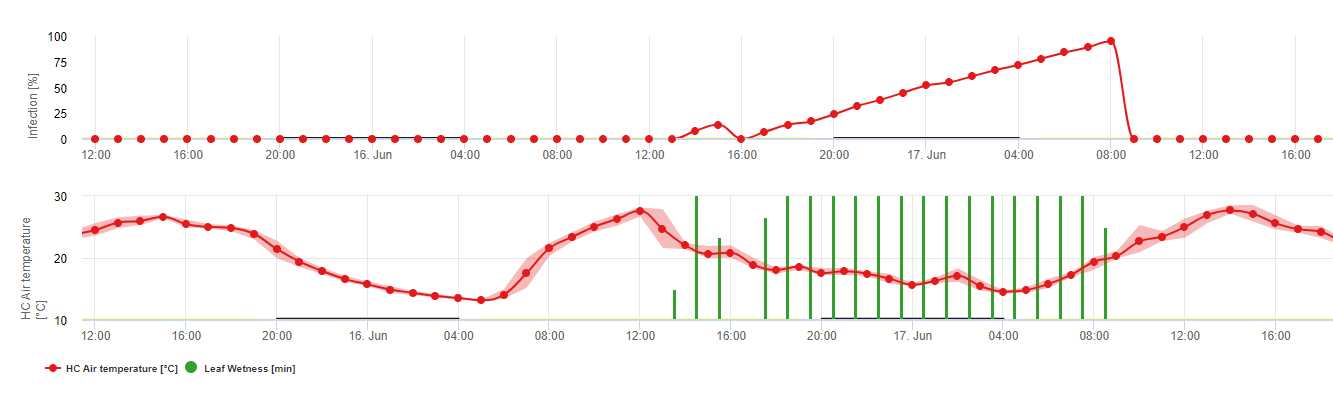
Infekce pocházející z oospor se nazývají primární infekce. Tento termín je zavádějící, protože na začátku léta se může objevit několik primárních infekcí. V epidemiologických událostech primární infekce nehrají důležitou roli, pokud je ve vinicích dostatek olejových skvrn a infekční potenciál letních spor (sporangií) převyšuje oospory.
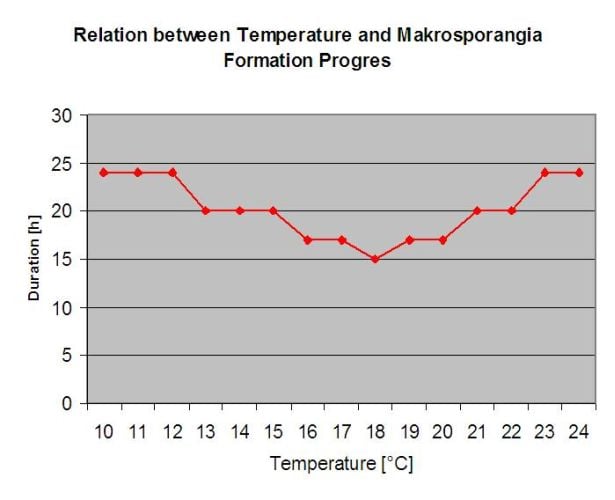
Přezimující oospory se vyvíjejí v tzv. makrosporangia vždy, když je po dobu přibližně 24 hodin dostatečná relativní vlhkost vzduchu. Makrosporangia uvolňují své zoospory do volné vody a silného deště (např. při bouřce). Spory jsou přenášeny na listy a/nebo zelené výhonky révy. Primární infekce tedy, potřebují delší deštivé období nebo několik po sobě jdoucích bouřek. První déšť způsobí, že opadané listy jsou nasyceny vodou, a silný déšť v následujících dnech způsobí, že makrosporangia uvolní své zoospory, které se dostanou na listy nebo výhonky révy. Pro úspěšnou infekci je nutná dostatečně dlouhá doba provlhčení listů, aby zoospory dosáhly stomu listů nebo výhonů a infikovaly je (na obrázku výše je znázorněn vývojový cyklus plísně révy vinné (Plasmopara viticola).
Model primární infekce padlím nejprve zjišťuje, zda je počasí vhodné pro vývoj makrosporangií. To platí, pokud jsou listy vlhké nebo pokud relativní vlhkost vzduchu po dešti neklesne pod 70%. V závislosti na teplotě se makrosporangia mohou vyvinout během 16 až 24 hodin. Pokud jsou k dispozici zralá makrosporangia, zobrazí se graficky na displeji primární infekce padlím. Pokud jsou makrosporangia přítomna, může silný déšť rozšířit jejich zoospory. Souvislý déšť o síle 5 mm je interpretován jako silný déšť a zoospory se šíří, primární infekce je než možná, pokud vlhkost listů trvala dostatečně dlouho.
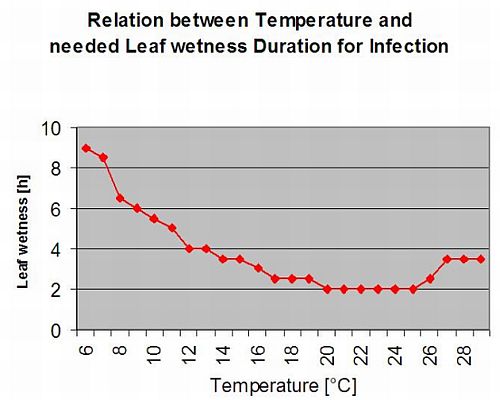
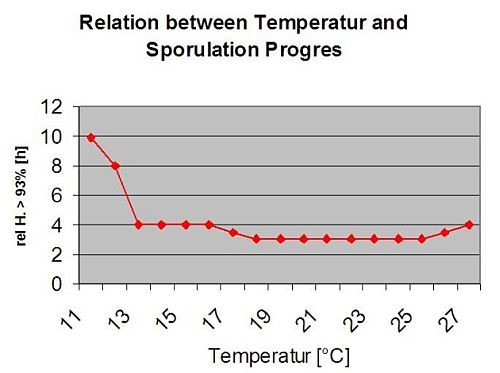
Sekundární infekce
Sekundární infekce Plasmopara viticola jsou možné pouze v případě, že na vinici nebo v její blízkosti existují již vyzrálá olejnatá místa. Olejové skvrny jsou zralé, když jsou schopny sporulovat (vytvářet sporangia). Sporangia jsou produkována pouze noc. Sluneční světlo inhibuje sporulaci. Sporangia se tvoří, pokud je teplejší než 12 °C a relativní vlhkost je velmi vysoká. Míra produkce sporangií se zvyšuje s teplotou až do 24 °C. Optimální teplota pro sporulaci na evropských odrůdách révy vinné (Vitis vinifera) je přibližně 24 °C. Pokud teplota překročí 29 °C, nemůže dojít ke sporulaci. V našem modelu zjišťujeme, zda vlhkost vyšší než 95% dochází během noci. Pokud tento stav trvá po dobu kumulativní hodinové teploty vyšší než 50 °C, sporulaci ukončí a nová sporangia Plasmopara existují na vinici. Například 50 °C hodin odpovídá 4 hodinám při 13 °C nebo 3 hodinám při 17 °C.
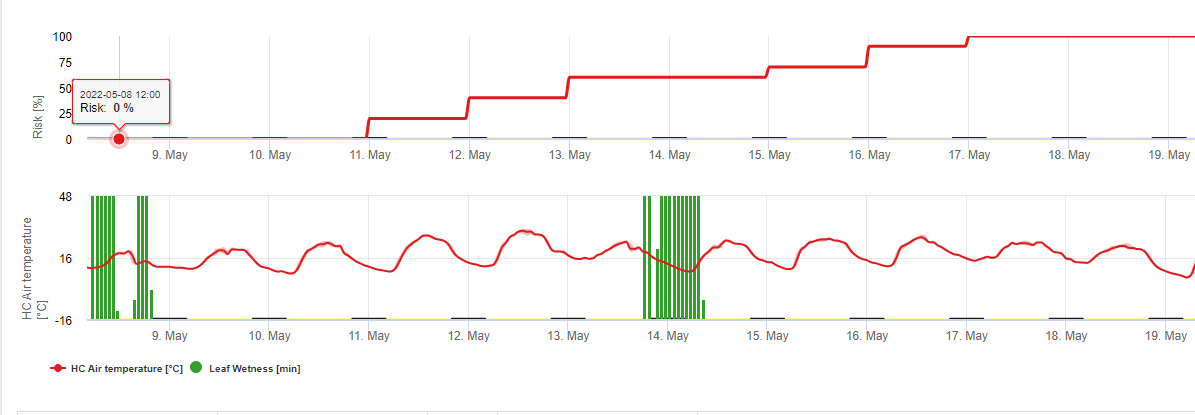
Následující graf znázorňuje vztah mezi teplotou a vlhkostí (relativní vlhkost, vlhkost listů, srážky), které se používají k modelování infekčních událostí v roce 2006. fieldclimate.com.
Moučnivku rajčat mohou celosvětově způsobovat tři patogeny.
Leveillula taurica (Oidiopsis taurica) je patogenem široké škály hostitelských druhů v teplém suchém až polosuchém podnebí v Asii, Středomoří, Africe a v poslední době i na jihozápadě Spojených států.
Erysiphe orontii (E. cichoracearum a E. polyphaga) je dalším druhem běžným na mnoha hostitelských rostlinách v mírném i tropickém pásmu.
A jako třetí druh Odium lycopersicum.
Faktory vzniku onemocnění
- relativní vlhkost > 50% (optimální relativní vlhkost > 90%).
- volná voda na povrchu listů není nutná
- teplotní rozsah: 10-35 °C (nejlépe pod 30 °C)
Prašná plíseň je choroba způsobená inokulem. Proto lze určit jen riziková období, hlavním faktorem poškození, např. vypuknutí epidemie, je počáteční inokulum (které je aktivní v širokém rozmezí teplot). Takže pro kontrolní strategie: kombinujte modelování rizikového období spolu s monitorováním houbového inokula (choroby) na poli!
Rajče na otevřeném poli
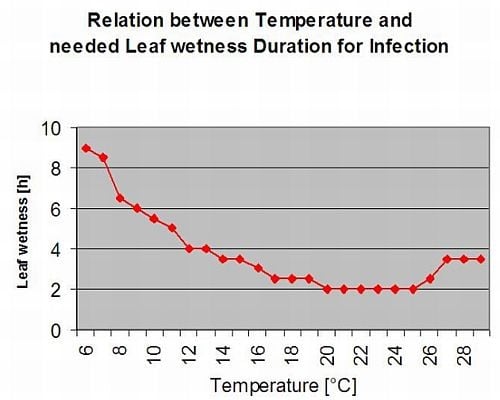
Rez chřestová Puccinia asparagi prospívá vlhké a teplé podnebí. Malé množství deště a vítr roznáší urediaspory, zatímco silné deště je mohou spláchnout do půdy. Z tohoto důvodu se období vlhka listů s více než 10 mm deště nebere jako období infekce. Vlhkost listů způsobená slabými dešti a rosou od 3 do 8 hodin při teplotách od 25 °C do 10 °C může být v létě a na konci léta obdobím infekce urediaspor.
Tento model odhaduje datum prvního výskytu C. beticola na základě kumulovaných průměrných denních teplot od prvního ledna nad 5 °C. V obdobích s relativní vlhkostí vyšší než 60% je prahová hodnota 1006,2 °C, a pokud bychom neměli žádnou relativní vlhkost, platí prahová hodnota 1081,9 °C.
Tento model byl vyvinut pro Itálii a používá se v Itálii a Německu.
Pro rok 2010 ve Štýrsku uvádí společnost CercoPrim 19. červen jako datum prvního postřiku. To je přibližně stejné datum, které by model rizika Pessl Instruments nebo model DIV určil pro náchylné odrůdy.
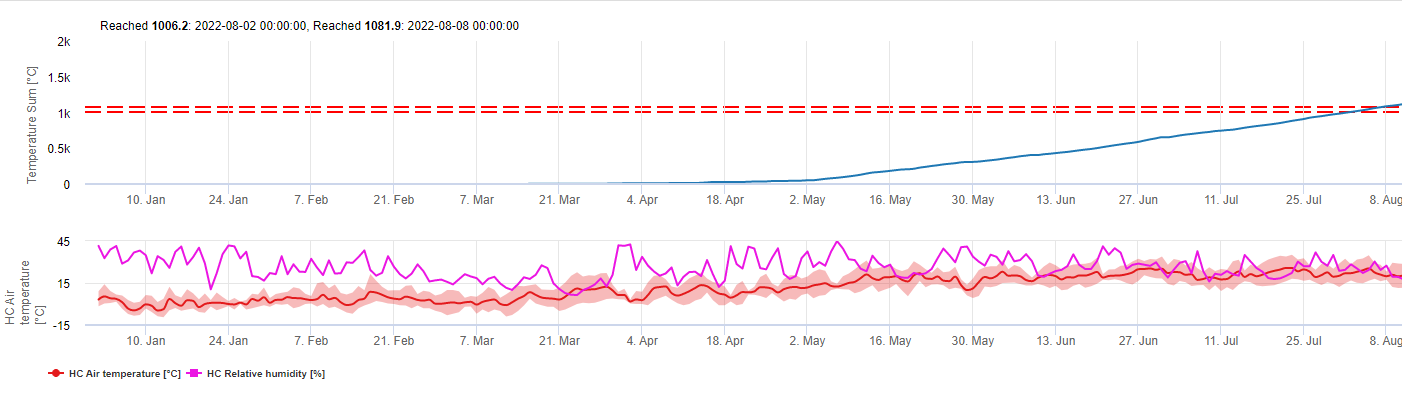
Patogen
Fialová skvrnitost chřestu a kapradí je způsobena houbou Stemphylium vesicarium. Houba přežívá zimu jako pohlavní spory (askospory) ve váčku (askus), který se vytváří v přezimujících strukturách (pseudothecia), jež se objevují jako malé černé tečky na zbytcích chřestu z předchozí sezóny. Na askospory se z askusu uvolňují za deště. a mohou být větrem přeneseny na nově vzešlé rostliny chřestu, kde způsobují primární infekci ve vegetačním období. Z těchto nových infekcí vznikají asexuálním procesem spory (konidie), které mohou způsobit sekundární infekce, což je proces, který se opakuje, dokud jsou teploty a srážky příznivé.
Nemoc
Výskyt fialové skvrnitosti jako významného problému při pěstování chřestu v Michiganu může být způsoben zavedením systému pěstování "no- till", při kterém se v dubnu poseče suché kapradí z předchozí sezóny a ponechá se na povrchu půdy. Tyto zbytky přetrvávají po celou dobu sklizně (polovina června) a jsou viditelné po celou dobu růstu kapradin (konec června až září) a jsou zdrojem askospor, které začínají infekci na počátku vegetačního období. Příznaky choroby jsou: Choroba se projevuje jako četné, mírně vpadlé, nafialovělé skvrny s hnědým středem, které se objevují na sklizených výhonech a kapradinách. Poškození na oštěpech se často vyskytují na návětrné straně, protože navátý písek způsobuje poranění, které podporuje infekci. V epidemických letech se skvrnitost může vyskytnout na 60-90% části kopí a může vést k odmítnutí úrody, zejména při prodeji čerstvého zboží. Skvrnitost se vyskytuje také na kapradinách chřestu, kde postihuje hlavní stonek, vedlejší větve a jehlice (kladofyly). Silná infekce kapradin může vést k předčasnému odlistění rostliny. Zvýšení intenzity výskytu fialové skvrnitosti je spojeno s delšími obdobími dešťů, mlhy nebo rosy.
Text: Mary K. Hausbeck, profesorka a odbornice na rozšiřování, Michiganská státní univerzita, katedra patologie rostlin, E. Lansing
TomCast pro chřest
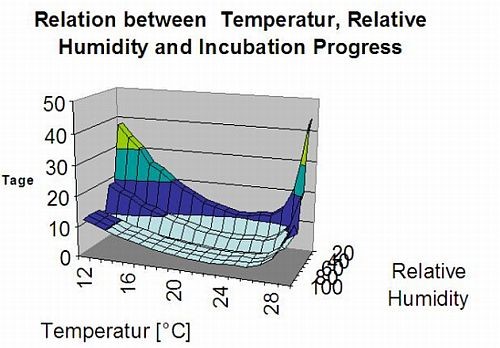
Souvislosti: TOMCAST (TOMato disease foreCASTing) je počítačový model založený na terénních datech, který se snaží předpovědět vývoj houbových chorob, a sice Raná skvrnitost, septoriová skvrnitost listů a antraknóza na rajčatech. Datalogery umístěné v terénu zaznamenávají hodinové údaje o vlhkosti listů a teplotě. Tyto údaje se analyzují po dobu 24 hodin a mohou vést k vytvoření hodnoty závažnosti choroby (DSV), což je v podstatě přírůstek rozvoje choroby. S narůstající hodnotou DSV se tlak choroby na plodinu dále zvyšuje. Pokud počet nahromaděných DSV překročí interval postřiku, doporučuje se aplikace fungicidu ke zmírnění tlaku choroby.
TOMCAST je odvozen z původního modelu F.A.S.T. (Forecasting Alternaria solani on Tomatoes), který vyvinuli doktoři Madden, Pennypacker a MacNab ? na Pennsylvania State University (PSU). Model F.A.S.T. z PSU byl dále upraven Dr. Pitbladem na Ridgetown College v Ontariu do podoby, kterou nyní známe jako model TOMCAST používaný Ohio State University Extension.
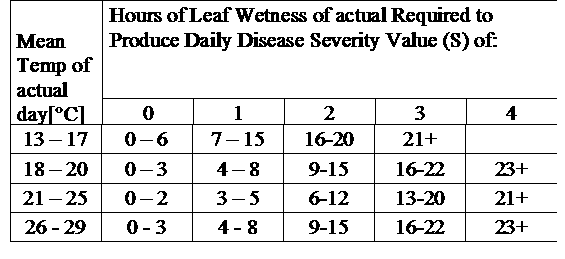
DSV DSV jsou: Hodnota závažnosti choroby (DSV) je měrná jednotka, která se udává pro určitý přírůstek rozvoje choroby (rané plísně).
Jinými slovy, DSV je číselné vyjádření toho, jak rychle nebo pomalu se choroba (raná plíseň) hromadí. DSV je určována dvěma faktory: vlhkostí listů a teplotou během "vlhkých listů". S rostoucím počtem hodin listové vlhkosti a teplotou se DSV hromadí rychleji. Viz níže uvedený graf závažnosti choroby.
Naopak při menším počtu vlhkých hodin na listech a nižší teplotě se DSV hromadí pomalu, pokud vůbec. Když celkový počet nahromaděných DSV překročí současnou hranici, která se nazývá interval postřiku nebo práh, doporučuje se postřik fungicidem, aby se ochránily listy a plody před rozvojem choroby.
Interval postřiku (který určuje, kdy byste měli postřik provést) se může pohybovat v rozmezí 15-20 DSV. Přesnou hodnotu DSV, kterou by měl pěstitel použít, obvykle dodává zpracovatel a závisí na kvalitě plodů a konečném využití rajčat. Dodržování intervalu postřiku 15 DSV představuje konzervativní použití systému TOMCAST, což znamená, že budete postřikovat častěji než pěstitel, který používá interval postřiku 19 DSV se systémem TOMCAST. Kompromis spočívá v počtu postřiků aplikovaných během sezóny a v potenciálním rozdílu v kvalitě plodů.
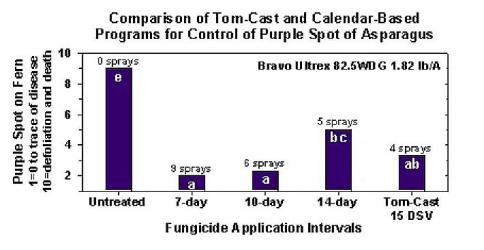
 TomCast s chřestem
TomCast s chřestem
Výzkum ukázal, že předpovědní přístroj Tom-Cast je slibnou alternativou ke kalendářním postřikům kapradin na komerčních polích chřestu. Tom-Cast upozorňuje pěstitele, když podmínky prostředí jsou příznivé pro rozvoj purpurové skvrnitosti. (delší období rosy nebo deště doprovázené vysokými teplotami). Účinné fungicidy aplikované podle předpovědi chorob Tom-Cast umožňují pěstitelům zvládnout fialovou skvrnitost chřestu a zároveň šetřit peníze a chránit životní prostředí.
Kredit za text: Jim Jasinski, koordinátor TOMCAST pro OHIO, INDIANA a MICHIGAN
Model infekce purpurovou skvrnou
Na stránkách Stemphylium versicarium model infekce vychází z práce SUHERIHO a PRICE v oblasti cibule a LLORENTA, VILARDELLA, BUGIANIHO, GHERARDIHO a MONTESINA v oblasti hrušek.
Jsou vypočteny křivky infekce pro lehkou, středně těžkou a těžkou infekci.
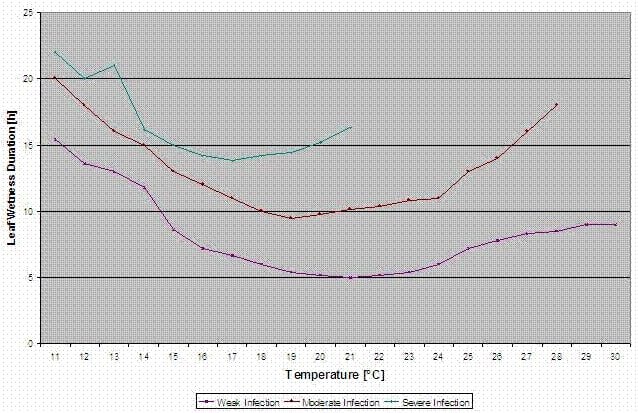
Při tomto rozdělení na příznivé, středně příznivé a velmi příznivé situace je na pěstiteli, aby rozhodl, jak velký bude tlak chorob na konkrétním poli a zda musí pokrýt konkrétní infekci.
Podmínky:
Teplota: 11-30°C
Vlhkost listů > 0 (počátek infekce) nebo rel. vlhkost > 90% (jen pokud již před výpočtem byla infekce a stále se drží)
lehká infekce (11-30 °C), středně těžká infekce (13-30 °C), těžká infekce (16-30 °C), řada s délkou trvání vlhkosti listů (viz výše).
Reset: pokud je rel. vlhkost nižší než 70%.