

Patogen
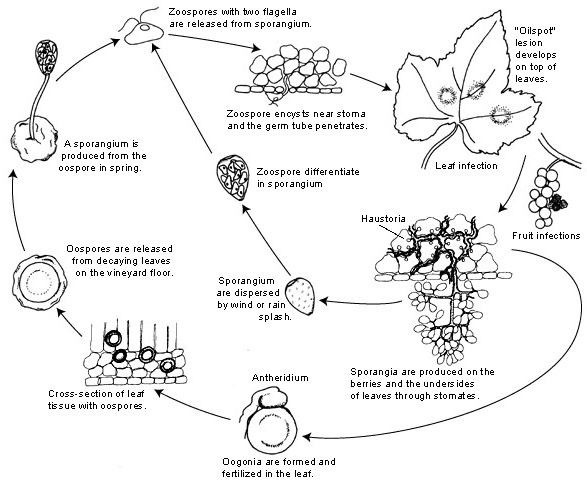
Patogenem plísně révy vinné je Plasmopara viticola. Je to obligátní parazit, což znamená, že k růstu potřebuje zelené a čerstvé orgány révy. Má asexuální i sexuální rozmnožovací stádia -zoospory a oospory. Existují tedy dvě stádia infekce: primární a sekundární infekce.
Primární infekce začíná klíčením přezimujících oospor, které na jaře za teplého počasí vytvářejí sporangia. Sporangia se vytvářejí v noci, protože sluneční světlo brání sporulaci. Silnostěnné oospory pomáhají patogenu přežít drsné zimní podmínky a zahájit primární infekci. Sporangia se šíří buď deštěm, nebo větrem a uvolňují zoospory ve volné vodě. Pro úspěšnou infekci je nutné dostatečně dlouhé zvlhčení listů nebo dešťové srážky.
Sekundární infekce je možná pouze v přítomnosti zralých olejových skvrn a probíhá prostřednictvím zoospor a sporangií. Zoospory a sporangia jsou velmi citlivé na nízkou vlhkost a světlo, což zkracuje jejich životaschopnost, což znamená, že k většině infekcí dochází brzy po uvolnění. Zoospory pronikají zárodečnými trubicemi do rostlinných pletiv a vyvíjejí hyfy pro další infekci. Tyto hyfy vytvářejí olejovité léze po inkubační době, která se pohybuje od 5 do 21 dnů v závislosti na teplotě - čím nižší teplota, tím déle to trvá.
Jakmile se opět ochladí, což je pro patogen nepříznivé, přejde k pohlavní tvorbě a produkuje oospory. Tyto oospory zůstávají během zimy v klidovém stavu a cyklus se opakuje.
Příznaky

Na horním povrchu listů se objevují drobné žluté léze, někdy obklopené hnědavou halou. Olejové skvrny se při dozrávání rozšiřují a integrují a haló mizí. Sporangia později vytvářejí na spodní straně lézí bílou skvrnitost a následuje nekróza. Olejnaté hnědé plochy se objevují také na výhoncích, stoncích a bobulích. Za teplých vlhkých nocí se mohou v důsledku sporulace pokrýt bílým chmýřím. Silná infekce způsobuje předčasný opad.
FieldClimate Modely
- V programu FieldClimate existují dva modely - primární a sekundární infekce.
- První graf zobrazuje inkubační dobu po ukončení infekce. Spodní grafy znázorňují průběh infekce pro slabou, střední a těžkou úroveň závažnosti. Infekci je třeba předpokládat, když rostoucí křivka v grafu dosáhne 100% - postřik lze zvážit, pokud se projeví závažná úroveň infekce.
Primární infekce
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
- Srážky
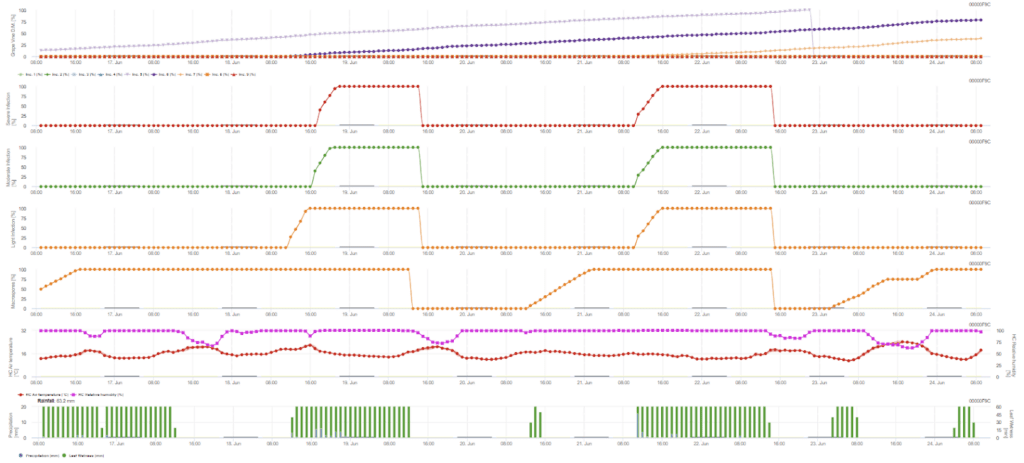
Primární infekce prověřuje, zda je počasí vhodné pro vývoj sporangií. To platí, pokud jsou listy vlhké nebo pokud relativní vlhkost vzduchu po dešti neklesne pod 70%. Sporangia se mohou vyvinout během 16 až 24 hodin v závislosti na teplotě. Souvislý déšť o síle 5 mm je interpretován jako silný déšť, který může šířit zoospory.
Sekundární infekce
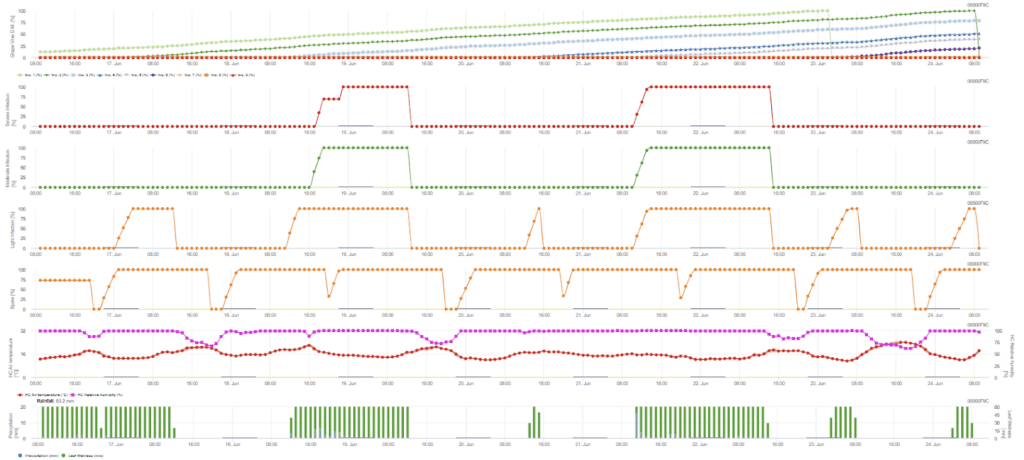
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
- Srážky
Sekundární infekce se kontroluje, pokud je teplota vyšší než 12 °C a relativní vlhkost vyšší než 95%. Rychlost produkce sporangií se zvyšuje při teplotách do 23 °C. Pokud tento stav trvá po dobu kumulativní hodinové teploty vyšší než 50 °C, předpokládáme, že sporulace je ukončena a ve vinici existují nová sporangia. Kumulovaných 50 °C odpovídá například 4 hodinám při 13 °C nebo 3 hodinám při 17 °C. Sporangia rychle odumírají, když se oteplí a vysuší - když relativní vlhkost klesne pod 50%, vynulujeme ji a když teplota překročí 29 °C, nemůže probíhat žádná sporulace.
Literatura
- Ash, G. (2000). Plíseň révy vinné. The Plant Health Instructor.
- Gessler, C., Pertot, I., & Perazzolli, M. (2011). Plasmopara viticola: přehled poznatků o plísni révy vinné a účinném managementu choroby. Phytopathologia Mediterranea, 50(1), 3-44.
- Kennelly, M. M., Gadoury, D. M., Wilcox, W. F., Magarey, P. A., & Seem, R. C. (2007). Primární infekce, produktivita lézí a přežívání sporangií u patogenu plísně révy vinné Plasmopara viticola. Phytopathology, 97(4), 512-522.
- Koledenkova, K., Esmaeel, Q., Jacquard, C., Nowak, J., Clément, C., & Ait Barka, E. (2022). Plasmopara viticola původce padlí révy vinné: od její taxonomie k managementu choroby. Frontiers in Microbiology, 13, 889472.
Patogen
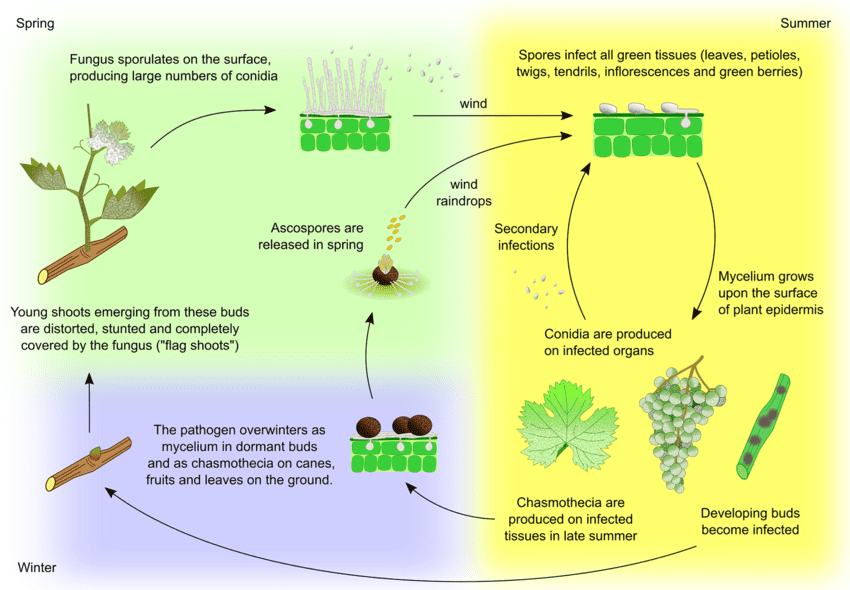
Původcem moučnatosti hroznů je Uncinula necator. Existují dva hlavní zdroje inokula - přezimující mycelium a askospory z kleistothecií - jejichž význam se může lišit v závislosti na oblasti.
Mycelium vytváří na jaře vlajkové výhonky. Vlajkové výhonky jsou částečně nebo zcela pokryté plísní a vznikají z latentně infikovaných pupenů. Protože se v nich tvoří kolonie, jsou tyto výhony dobře viditelné a často mají deformované listy.
Cleistothecia uvolňují askospory brzy na jaře. Důležitým faktorem pro uvolňování askospor jsou dešťové srážky. Se stoupající teplotou dochází k dehiscenci častěji. Askospory klíčí do 12 hodin, vytvářejí zárodečné trubice a apresorium na rostlinných pletivech, což vede k rozptýlené tvorbě kolonií.
K sekundární infekci dochází prostřednictvím konidií produkovaných v koloniích. Stejně jako askospory klíčí a vyvíjejí se z nich zárodečné trubice a apresorium. U. necator nepotřebuje k infekci volnou vodu a k tvorbě konidií vysokou relativní vlhkost. Když se povětrnostní podmínky stanou méně příznivými, houba vytváří kleistothecia. Přezimuje buď v kleistotheciích, nebo jako vegetativní mycelium v dormantních infikovaných pupenech a cyklus se opakuje.
Příznaky

Příznaky se vyskytují na všech zelených částech révy. Na výhonech se vytvářejí kolonie a pokrývá je bílošedé mycelium, tzv. "vlajkové výhonky". Z nich vyrůstají deformované listy. Kolonie plísně se vyskytují také na listech - buď na spodní, nebo na obou stranách. Na bobulích se objevují popelavě šedé skvrny a v závažných případech se rozštěpují. Na třapinách se objevují tmavě červenohnědé skvrny a mohou odumírat od špiček.
FieldClimate Modely
Infekce askosporami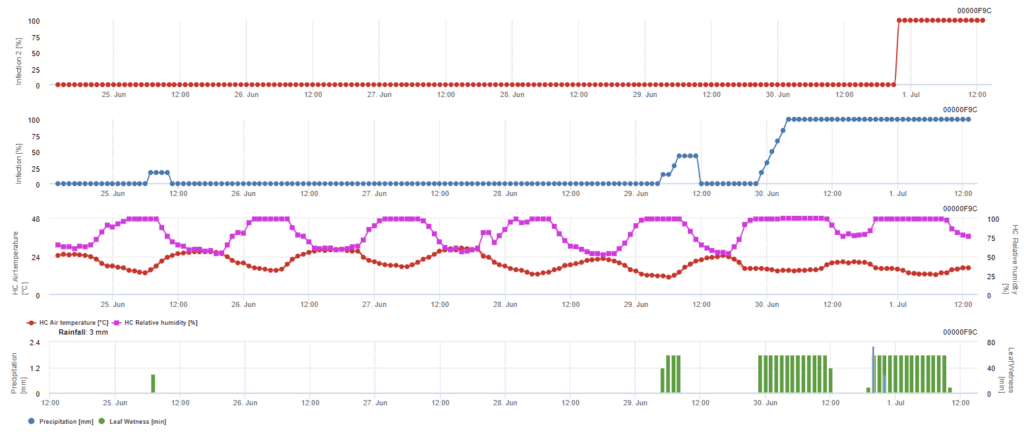
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
- Srážky
Tento model se doporučuje v případě, že se askospory tvoří v chladném zimním období (nedochází tedy k infekci myceliem). Infekce askosporami předpovídá uvolnění askospor a počáteční infekci na základě průměrných teplot během delšího období vlhkosti listů. Pro výpočet je k uvolnění askospor zapotřebí přibližně 2,5 mm srážek, po nichž následuje minimálně 8 až 12 hodin vlhkosti listů a teploty mezi 10 až 15 °C. Jakmile dojde k infekci, model přechází do fáze hodnocení rizika choroby (model Californian Risk) a je založen na vlivu teploty na rychlost reprodukce patogenu.
Kalifornský rizikový model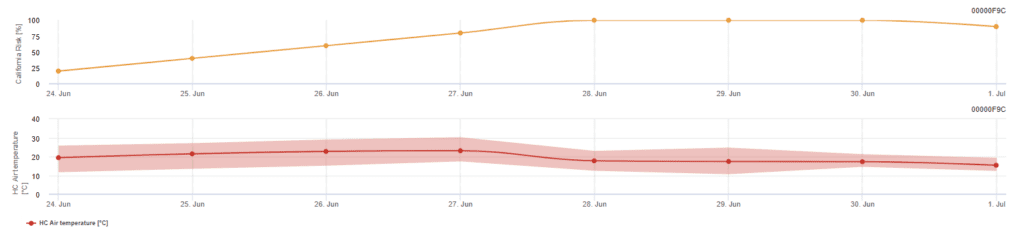
Potřebné senzory:
- Teplota vzduchu
Model vychází z laboratorních studií v Kalifornii. Po uvolnění askospor a vyklíčení (model) jsou následný vývoj a rozmnožování plísně ovlivněny teplotami. Vyhodnocuje teploty a posuzuje riziko rozvoje moučnaté plísně pomocí indexu 0 až 100 bodů.
Pro zahájení indexu hodnocení rizika jsou nutné tři dny v řadě s teplotou v rozmezí 21 až 30 °C minimálně šest hodin po sobě. Ten získává 20 bodů za každý den, který splňuje 6 nebo více po sobě jdoucích hodin mezi 21 a 32 °C, a ztrácí 10 bodů za ty, které je nesplňují, nebo když teplota překročí 32 °C nebo klesne pod 21 °C.
Nízké hodnoty indexu 0~30 naznačují, že se patogen nerozmnožuje. Index 40~50 je považován za středně vysoký a znamená, že se plíseň rozmnožuje přibližně 15 dní. Hodnoty indexu nad 60 znamenají, že se patogen rozmnožuje rychle (každých 5 dní) a že riziko epidemie choroby je velké.
Rizikový model Pessl Instruments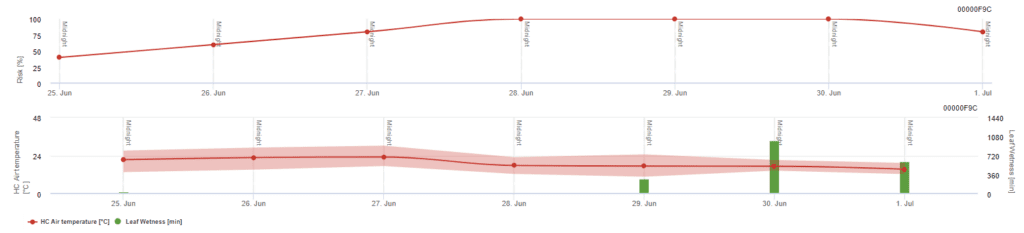
Potřebné senzory:
- Teplota vzduchu
- Vlhkost listů
Vedle teploty vzduchu může být řídícím faktorem i vlhkost listů, protože dlouhá období vlhkosti listů vedou k usídlení antagonistické houby (Ampelomyces quisqualis), která vede k poklesu výskytu U. necator. Základy modelu jsou ekvivalentní modelu Californian Risk, ale v tomto modelu je zohledněna také vlhkost listů. Vlhkost listů delší než 8 hodin vede k poklesu o 10 bodů.
Nízké hodnoty indexu 0~20 naznačují, že se patogen nerozmnožuje. Index 20~60 je považován za mírný a platí pro něj běžný interval postřiku. Hodnoty indexu nad 60 znamenají, že se patogen rychle množí (každých 5 dní) a že riziko epidemie choroby je velké, takže se doporučuje zkrátit interval postřiku.
Literatura
- Gadoury, D. M., & Pearson, R. C. (1990). Ascocarp dehiscence and ascospore discharge in Uncinula necator (Dehiscence askokarpu a výtok askospor u Uncinula necator). Phytopathology, 80(4), 393-401.
- Gadoury, D. M., & Pearson, R. C. (1990). Germination of ascospores and infection of Vitis by Uncinula necator [Klíčení askospor a infekce Vitis houbou Uncinula necator]. Phytopathology, 80(11), 1198-1203.
- Hall, T. W. (2000). Epidemiologie plísně révy vinné, Uncinula necator, ve Willamette Valley.
- Halleen, F., & Holz, G. (2001). An Overview of the Biology, Epidemiology and Control of Uncinula Necator (Powdery Mildew) on Grapevine, with Ref Ere Nee to South Africa (Přehled biologie, epidemiologie a kontroly plísně Uncinula Necator na révě vinné s odkazem na Jižní Afriku). South african journal of Enology and Viticulture, 22(2), 111-121.
- Rügner, A., Rumbolz, J., Huber, B., Bleyer, G., Gisi, U., Kassemeyer, H. H., & Guggenheim, R. (2002). Tvorba přezimovacích struktur Uncinula necator a kolonizace révy vinné v polních podmínkách. Plant pathology, 51(3), 322-330.
Patogen
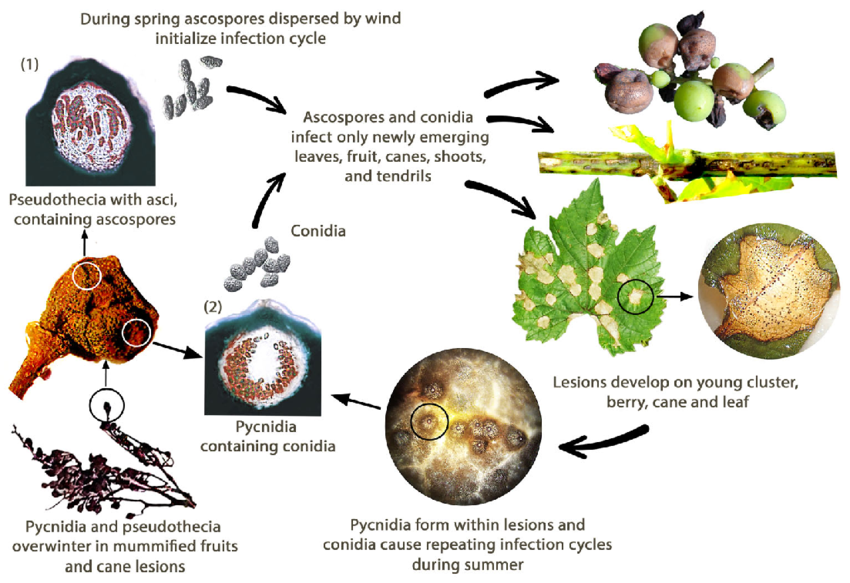
Černou hnilobu révy vinné způsobuje houba Guignardia bidwellii. Houba přezimuje v různých částech vinné révy a může přežívat více než dva roky v lézích napadených výhonů.
Na jaře produkují pyknidy konidie (nepohlavní spory) a pseudothecia askospory (pohlavní spory). Konidie se obvykle šíří na krátké vzdálenosti, zatímco askospory mohou infikovat na větší vzdálenosti. Tyto spory jsou přenášeny větrem a deštěm, aby infikovaly mladé tkáně vnímavých hostitelů a iniciovaly primární infekci. Konidie slouží také jako sekundární inokulum, což přispívá k rychlému a opakovanému šíření choroby.
Během srpna se pyknidy přemění v přezimující stadium, které produkuje pseudothecia, z nichž se vytvářejí askospory. Tyto askospory jsou důležitým zdrojem primárních infekcí na jaře následujícího roku.
Příznaky

Vznikají červenohnědé skvrny, které se nakonec spojí a často jsou obklopeny malými černými tečkami, což jsou houbové struktury známé jako pyknidy, které produkují konidie. K této infekci jsou náchylnější zejména mladé a rychle rostoucí listy.
K infekci plodů dochází obvykle po opadnutí kalichu, přičemž většina příznaků se projeví, když jsou plody poloviční až téměř plné. Zpočátku se objevují malé skvrny obklopené hnědým prstencem, které se pak zvětšují a tmavnou, až nakonec s rozvojem choroby pokryjí celou bobuli. Během několika dnů mohou napadené plody mumifikovat, roztříštit se nebo předčasně opadat.
Kromě toho jsou postiženy i další části rostlin, jako jsou výhonky, stonky a úponky. Vznikají oválné, fialové až černé léze, na jejichž povrchu jsou rozptýleny pyknidy.
FieldClimate Model
Vinařství Černá hniloba Model
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
- Srážky
Model původně vychází z literatury publikované Spottsem, ale s úpravami Daniela Molitora. Revize zavedla tři třídy závažnosti (lehká, středně těžká a těžká) podobně jako u modelů strupovitosti jablek. Při vysokých teplotách je doba vlhnutí listů 8 hodin dostatečná k tomu, aby byla infekce příznivá, a předpokládá se, že infekce je ukončena, když graf dosáhne 100%.
Závažnost infekce závisí na době trvání optimální teploty a vlhkosti houby. Infekce splňující Spottsova kritéria jsou hodnoceny jako lehké infekce. Infekce splňující Spottova kritéria u 150% a 200% jsou hodnoceny jako středně těžké, resp. těžké infekce.
V závislosti na infekčním tlaku je třeba infekce preventivně zakrýt nebo krátce po infekci aplikovat léčebný postřik. V mírném polosuchém klimatu Mosely nebo většiny rakouských vinařských oblastí by se postřik po zjištění lehké infekce nedoporučoval.
Literatura
- Wilcox, Wayne F. "Černá hniloba Guignardia bidwellii." Identifikační list choroby č. 102GFSG-D4. 2003. Cornell. 24. 10. 2010
- http://www.nysipm.cornell.edu/factsheets/grapes/diseases/grape_br.pdf
- Ellis, Michael A. "Informační list Zemědělské a přírodní zdroje: "Černá hniloba hroznů." Oddělení patologie rostlin. The University of Ohio State Extension. 2008
- http://ohioline.osu.edu/hyg-fact/3000/pdf/HYG_3004_08.pdf
- Molitor, D. (2009). Untersuchungen zur Biologie und Bekämpfung der Schwarzfäule (Guignardia bidwellii) an Weinreben. Gesellschaft zur Förderung der Forschungsanstalt Geisenheim.
- Ries, S. M. (1999). Zprávy o chorobách rostlin: Sborník zpráv o chorobách révy vinné: Černá hniloba hroznů. Integrated Pest Management at the Uni. of Illinois. http://ipm. illinois. edu/diseases/series700/rpd703.
- Spotts, R. A. (1977). Vliv délky trvání vlhkosti listů a teploty na infekčnost Guignardia bidwellii na listech vinné révy. Phytopathology, 67(11), 1378-1381.
- Szabó, M., Csikász-Krizsics, A., Dula, T., Farkas, E., Roznik, D., Kozma, P., & Deák, T. (2023). Černá hniloba hroznů (Guignardia bidwellii) - komplexní přehled. Horticulturae, 9(2), 130.
- http://extension.cropsciences.illinois.edu/fruitveg/pdfs/771-BlackRotOfGrape.pdf
- https://www.missouribotanicalgarden.org/gardens-gardening/your-garden/help-for-the-home-gardener/advice-tips-resources/insects-pests-and-problems/diseases/fruit-spots/black-rot-of-grapes
Patogen

Lobesia botrana v závislosti na regionu obvykle dokončí dvě až tři generace za rok. První generace (květen a červen) napadá pouze květy, takže ošetření je nutné pouze v případě, že je populace molů obzvláště vysoká. Druhá generace (červenec a srpen) a třetí generace způsobují největší škody, přičemž třetí generace je obzvláště škodlivá, protože se časově shoduje s dozráváním hroznů. Čtvrtá generace se může vyskytnout v teplejším podnebí, ale ošetření není zpravidla nutné, protože se časově shoduje se sklizní hroznů.
Můra přezimuje v kuklách v zámotcích, a když na jaře stoupnou teploty, objeví se dospělci první generace, přičemž samci se obvykle objeví dříve než samice. Let první generace začíná v době, kdy praskají pupeny, a trvá 4 až 5 týdnů, během nichž dochází k páření. Po jednom až dvou dnech po páření kladou samičky 80 až 160 vajíček.
Vajíčka o průměru 0,6 až 0,8 mm jsou viditelná pouhým okem. Zpočátku jsou krémově bílá, s vývojem zárodku se mění na žlutá, s černou barvou hlavy larvy. Vajíčka se líhnou po 66 stupních Celsia (DDC), tedy přibližně po 7 až 11 dnech. Larvy společně pavučinkují části květů, přičemž jejich vývoj trvá 20 až 30 dní. Následuje vylíhnutí a dospělci se objevují o 6 až 14 dní později.
"Stupeň-den" se vypočítá tak, že se změří, o kolik průměrná denní teplota překročí určitou základní mezní teplotu potřebnou pro vývoj. S rostoucí teplotou nad základní hranicí se počet stupňů za den zvyšuje.
Dospělé můry jsou asi 6 až 8 mm dlouhé a rozpětí křídel je 11 až 13 mm. Samičky jsou o něco větší než samečci, i když obě mají podobně mozaikovitě vzorovaná křídla.
Vajíčka druhé a třetí generace se líhnou rychleji, během 3 až 5 dnů. Samičky kladou vajíčka jednotlivě na zastíněné bobule a po vylíhnutí larvy vnikají do bobulí a vydlabávají je. Na podzim vyvolávají noci delší než 11 hodin diapauzu, klidové stadium, které kuklám umožňuje lépe snášet chlad a přezimovat.
Příznaky
Housenky první generace se živí na květenstvích révy, vydlabávají květní pupeny a otáčejí je, aby je ochránily, což vede k poškození uvnitř hroznu. Ve druhé generaci se larvy zaměřují na vyvíjející se bobule, čímž vytvářejí vstupní místa pro houbové patogeny, jako jsou např. Botrytis cinerea. Toto přímé krmení způsobuje viditelné škody, které se obvykle projevují tmavými skvrnami v okolí míst krmení. Největší škody způsobuje třetí generace, kdy larvy pronikají do dozrávajících bobulí a živí se jimi.
Zůstávají po nich scvrklé bobule a výkaly larev. Útržky epidermální tkáně bobulí zůstávají volně přichyceny ke stopkám spolu se suchou, vydlabanou "slupkou" bobulí.
FieldClimate Model
Grapevine Berry Moth Model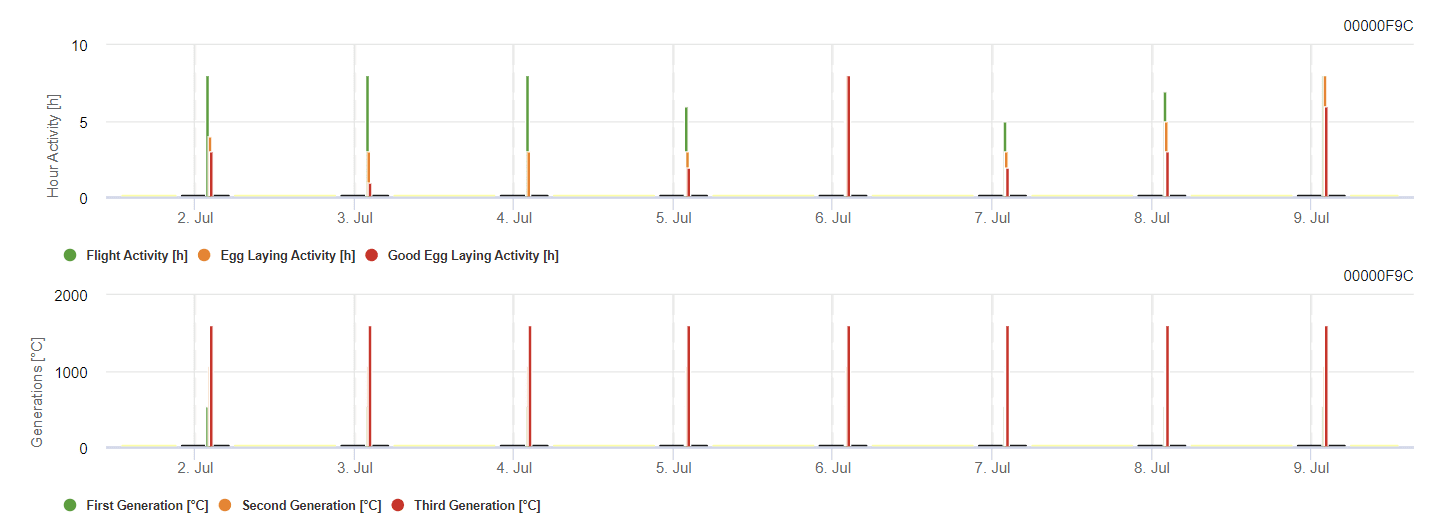
Potřebný senzor:
- Teplota vzduchu
Model vypočítává riziko podle teploty vzduchu. První graf ukazuje období letové aktivity s kladením vajíček a dobrou aktivitou hmyzu při kladení vajíček. Druhý graf ukazuje, jakou generaci je třeba v daném období očekávat. Ve třetím grafu jsou zobrazeny klimatické údaje, které ukazují kumulaci stupňových dnů (teploty > 8 °C až 24 °C za hodinu děleno 24).
Literatura
- Varela, L. G., Smith, R. J., Cooper, M. L., & Hoenisch, R. W. (2010). European grapevine moth, Lobesia botrana. Vinice v údolí Napa Valley. Pract. Winery Vineyard, 2010, 1-5.
- https://ipm.ucanr.edu/invasive-and-exotic-pests/european-grapevine-moth/
- https://www.ages.at/en/plant/plant-health/pests-from-a-to-z/grape-berry-moth
Patogen
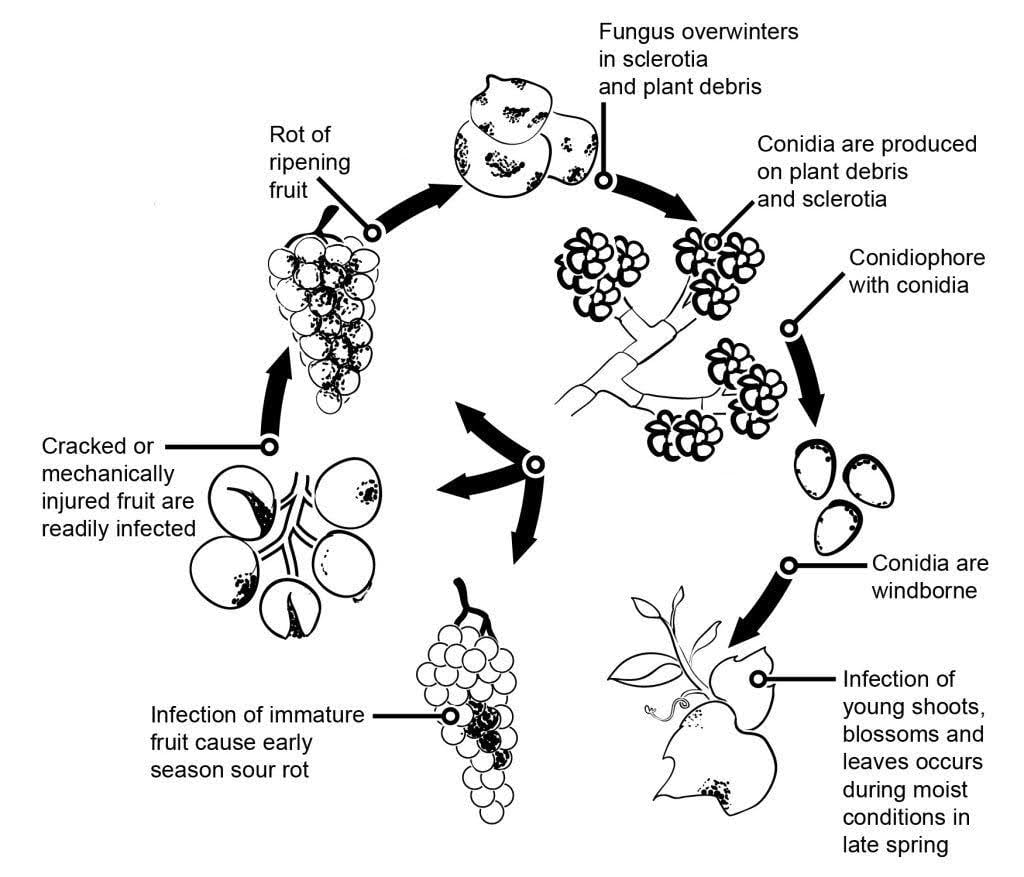
Původcem plísně šedé na révě vinné je Botrytis cinerea.
Epidemiologii plísně šedé významně ovlivňují latentní infekce iniciované infekcí květu. Byly identifikovány různé cesty od květu k plodu: houba infikuje květní stopku a dostane se až do vajíčka, kde zůstává latentní díky předem vytvořenému obrannému mechanismu rostliny. Rozhodující jsou také infekce přes tyčinky, okvětní lístky a kališní lístky. B. cinerea může infikovat tyčinky a systematicky růst směrem k čnělce a šířit se do stopky a cévních pletiv v bobulích. Další cesty zahrnují také saprofytický růst patogenu.
Primárním zdrojem infekce na jaře jsou přezimující mycelia nebo sklerocia. Konidie (nepohlavní spory) jsou produkovány a rozptylovány větrem a deštěm. Po dopadu konidie klíčí a vytvářejí zárodečné trubice a apresoria, která pronikají do rostliny a infikují ji. Houba může zůstat latentní, dokud plody nedozrají a nezvýší se obsah cukru. Konidie vznikající z primárních zdrojů inokula se řídí denním cyklem iniciace, produkce a šíření.
Zatímco pohlavní stadium B. cinerea se vyskytuje jen zřídka, patogen se stává aktivnějším, když bobule dozrávají. Zvýšený obsah cukru v bobulích zvyšuje náchylnost k infekci. Stejně tak jsou květy vysoce náchylné, když opadávají, přičemž hojný pyl zvyšuje závažnost infekce.
Příznaky

Plíseň šedá napadá především zralé bobule hroznů, které se zpočátku jeví jako měkké a nasáklé vodou. Časem se bobule zbarví do červenohněda a seschnou. Za příznivých podmínek se pokryjí šedou hmotou houbového mycelia a konidií. Zdravé bobule se mohou nakazit i přímým kontaktem s nemocnými.
Houba může také způsobit plíseň květů, která může vést ke značným ztrátám na úrodě na počátku sezóny. Ačkoli jsou infekce listů vzácné, pokud se objeví, začínají jako matné zelené skvrny podél žilek, které se nakonec vyvinou v nekrotické léze.
FieldClimate Model
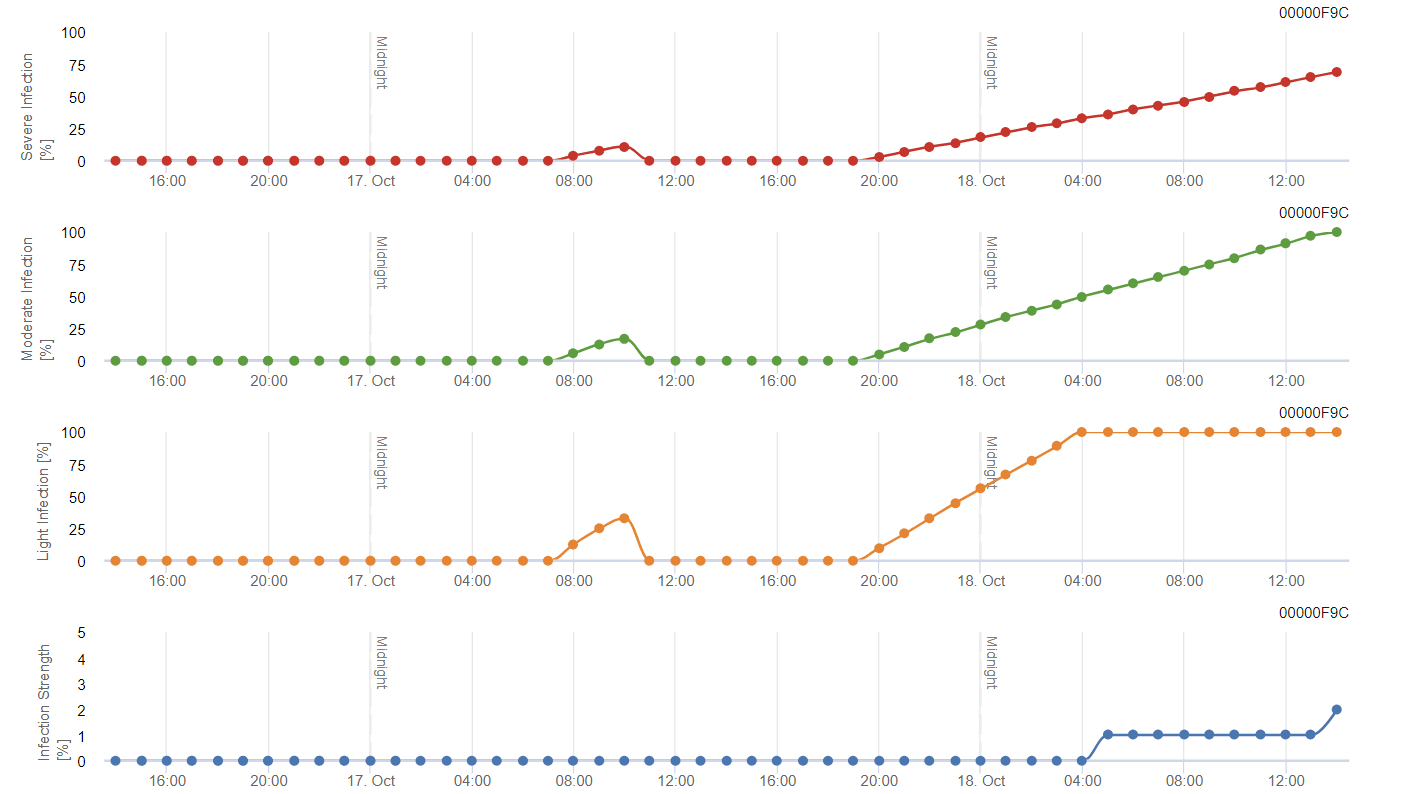
Obecný model šedé formy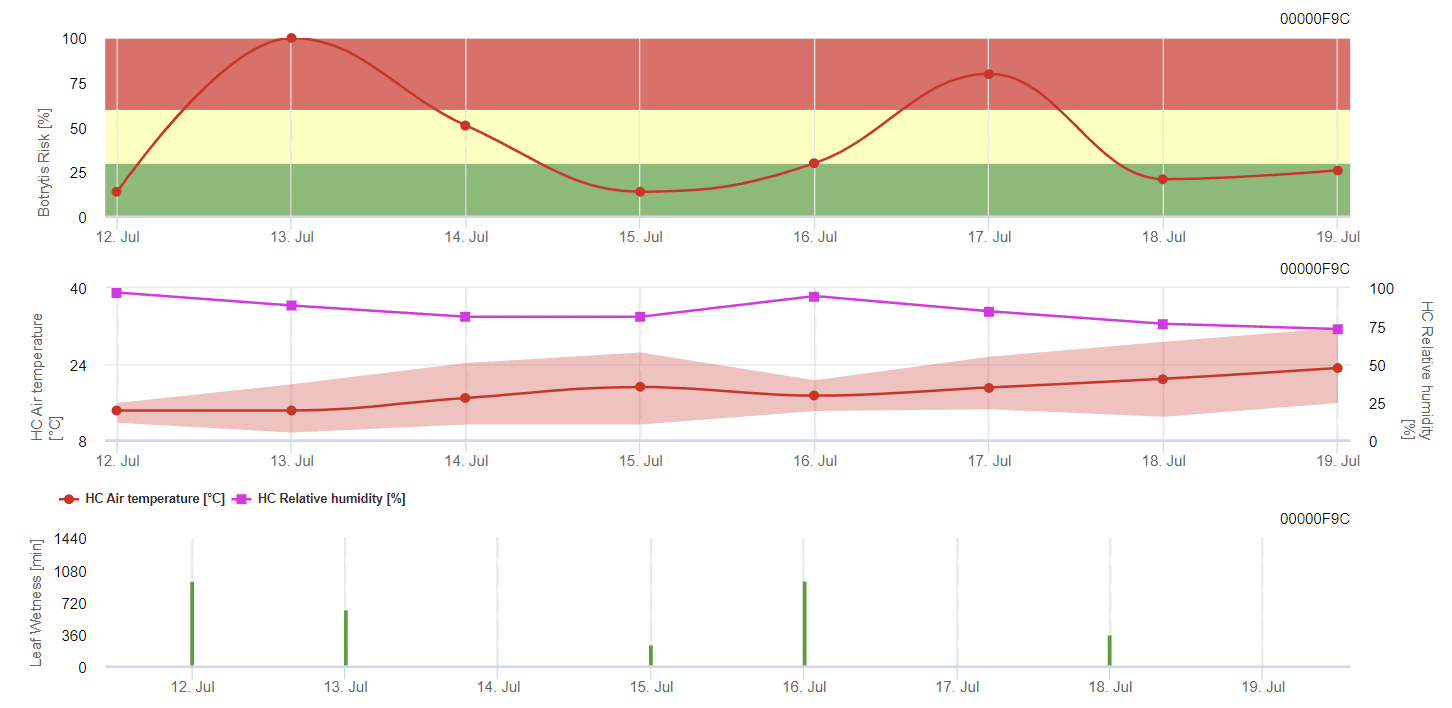
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
B. cinerea souvisí s vlhkým klimatem. Houba vyžaduje k infekci vysokou relativní vlhkost nebo přítomnost volné vody. Teplá období s delší dobou vlhkosti listů vedou ke zvýšenému riziku výskytu B. cinerea, zatímco suchá období ho snižují. K infekci dochází na mladých výhonech během dlouhých vlhkých období nebo škodlivých bouří s krupobitím.
Model počítá riziko v rozmezí hodnot 0 až 100%. Tato hodnota udává tlak B. cinerea v daném čase - pokud máme hodnotu 100%, znamená to, že několikrát došlo k dostatečně dlouhému zvlhčení, aby došlo k infikování vnímavé tkáně. Počítáme tzv. mokré body mezi vlhkostí listů a teplotou s maximem původně 38400 bodů (začátek sezóny, který vykazuje riziko 30%). Po tomto období každé vlhké období s přibližně 4000 vlhkými body zvyšuje riziko o 10%, nebo naopak každé suché období snižuje riziko o ⅕ původní hodnoty.
Literatura
- Broome, J. C., English, J. T., Marois, J. J., Latorre, B. A., & Aviles, J. C. (1995). Development of an infection model for Botrytis bunch rot of grapes based on wetness duration and temperature (Vývoj modelu infekce Botrytisovou hnilobou hroznů na základě délky trvání vlhkosti a teploty). Phytopathology, 85(1), 97-102.
- Elmer, P. A., & Michailides, T. J. (2007). Epidemiologie Botrytis cinerea v ovocných sadech a vinné révě. In Botrytis: Biology, Pathology and Control (s. 243-272). Dordrecht: Springer Netherlands.
- Williamson, B., Tudzynski, B., Tudzynski, P., & Van Kan, J. A. (2007). Botrytis cinerea: příčina onemocnění plísní šedou. Molecular plant pathology, 8(5), 561-580.
- Ciliberti, N., Fermaud, M., Roudet, J., & Rossi, V. (2015). Podmínky prostředí ovlivňují infekci zralých bobulí révy vinné Botrytis cinerea více než kmen nebo genotyp transpozonu. Phytopathology, 105(8), 1090-1096.
Patogen
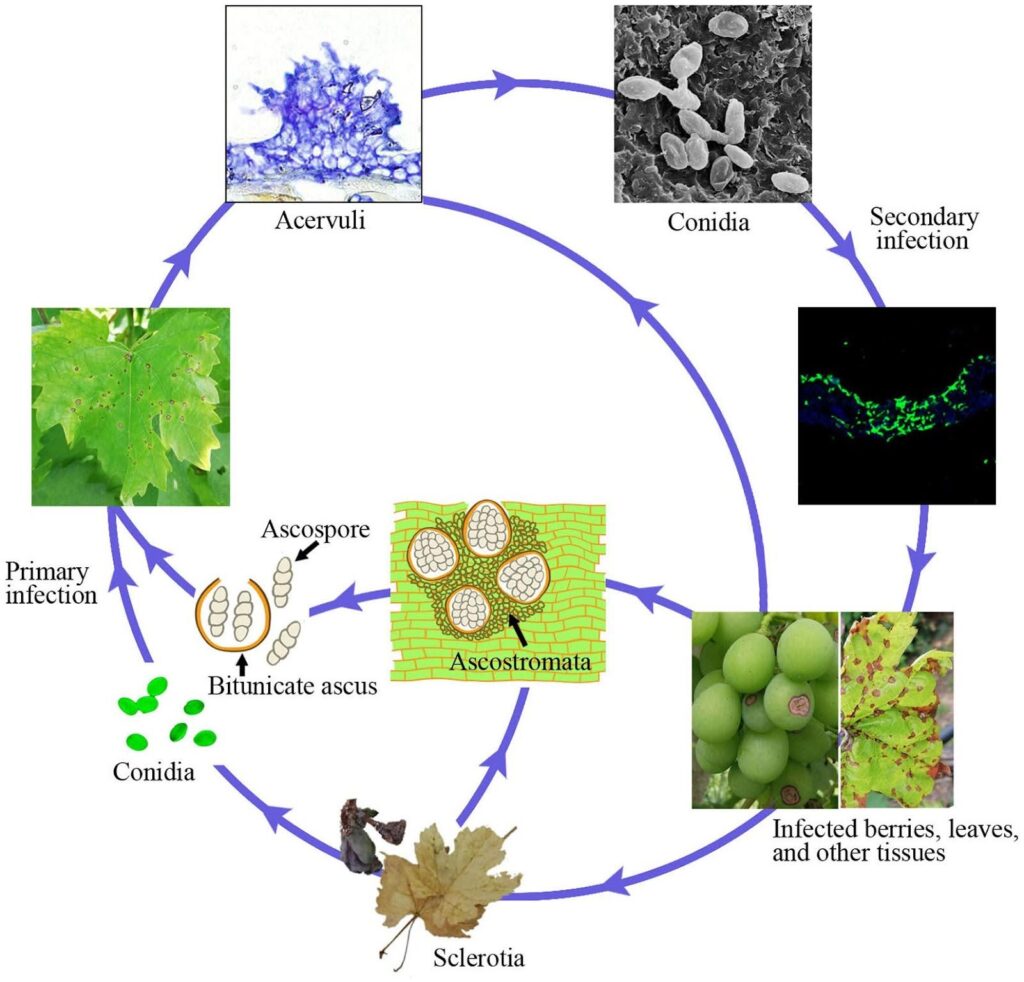
Antraknózu révy vinné způsobuje patogen Elsinoe ampelina.
Hlavním zdrojem nákazy jsou infikované hůlky. Sklerocia a mycelia přežívající v zimě v lézích a bobulích jsou na jaře aktivní a produkují askospory (pohlavní spory) a konidie (nepohlavní spory) za vlhkých podmínek (déšť nebo rosa po dobu 24 hodin) v rozmezí teplot 2 ~ 40 °C.
Tyto spory jsou deštěm a větrem rozptýleny do nových tkání a po přistání klíčí a vytvářejí zárodečné trubice a apresoria, čímž iniciují novou infekci. Mohou infikovat nové listy, výhonky, úponky a mladé plody. Teplé počasí zkracuje dobu trvání vlhka potřebnou pro počáteční infekci a inkubační dobu. K optimálnímu klíčení spor dochází při teplotě 25 ~ 30 °C, přičemž je zapotřebí minimálně 3 ~ 4 hodiny vlhka v listech. Při infekci při teplotě kolem 10 °C je inkubační doba přibližně 14 dní.
S postupující kolonizací se objevují acervuly a vznikají nové konidie, které slouží jako sekundární infekční inokulum. Jsou zodpovědná za rychlou a další infekci v sezóně.
Příznaky
E. ampelina napadá nadzemní, šťavnaté části révy, včetně výhonů, listů, řapíků, úponků, řapíků a bobulí; nejčastěji dochází k poškození výhonů a bobulí. Hrozny mají ontogenní rezistenci vůči houbám, takže mladá pletiva jsou náchylnější.
Zpočátku se objevují malé červenohnědé skvrny, které se za vlhkého počasí zvětšují a stávají se mírně vpadlými s šedým středem obklopeným tmavým okrajem. Někdy mohou příznaky vypadat jako poranění způsobené krupobitím. Nekrotický střed na listech obvykle vypadává a vytváří vzhled "vystřelené díry".
Při silné infekci mohou rostliny vykazovat předčasnou defoliaci, zpomalení růstu třtiny, lámání stonků, opad bobulí a opožděný vývoj a dozrávání bobulí.
FieldClimate Model
Vinařství Model antraknózy
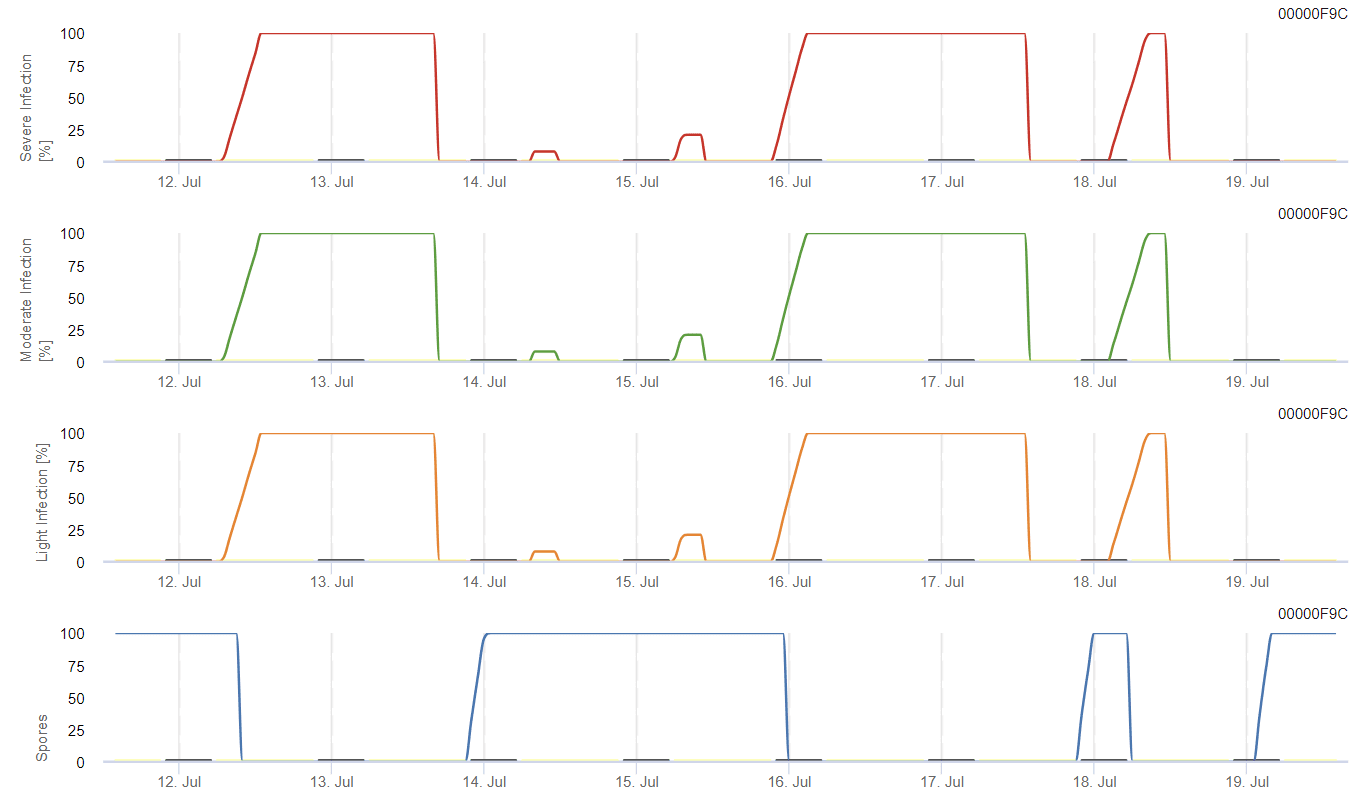
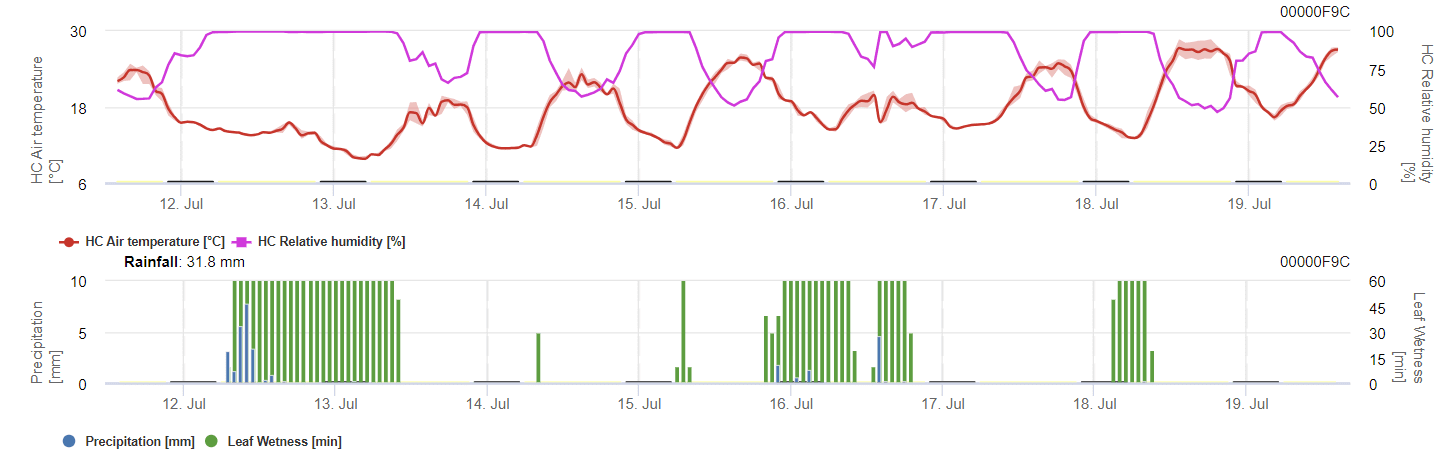
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
- Srážky
Model vypočítává riziko antraknózy na základě teploty vzduchu, relativní vlhkosti, vlhkosti listů a srážek.
Možný vývoj slabé, středně těžké a těžké infekce je zobrazen v samostatných grafech. Je třeba předpokládat, že infekce je dokončena, když rostoucí křivka dosáhne 100%. Poslední graf ukazuje sporulaci E. ampelina; pokud hodnota dosáhne 100%, předpokládá se, že spóry E. ampelina jsou přítomny.
Pro vývoj přezimujících spor by měly podmínky splňovat rozmezí teplot 2 až 40 °C, relativní vlhkost vyšší než 90% nebo vlhkost listů. Jakmile vývoj spor dosáhne 100%, začíná se počítat s infekcí. Závažnost infekce závisí na vlhkých podmínkách (dešťové události). Pokud však vlhkost klesne pod 50%, přestane se počítat jak vývoj spor, tak infekce.
Literatura
- Li, Z., Dos Santos, R. F., Gao, L., Chang, P., & Wang, X. (2021). Současný stav a budoucí perspektivy antraknózy révy vinné způsobené Elsinoe ampelina: An important disease in humid grape-growing regions. Molecular Plant Pathology, 22(8), 899-910.
Patogen
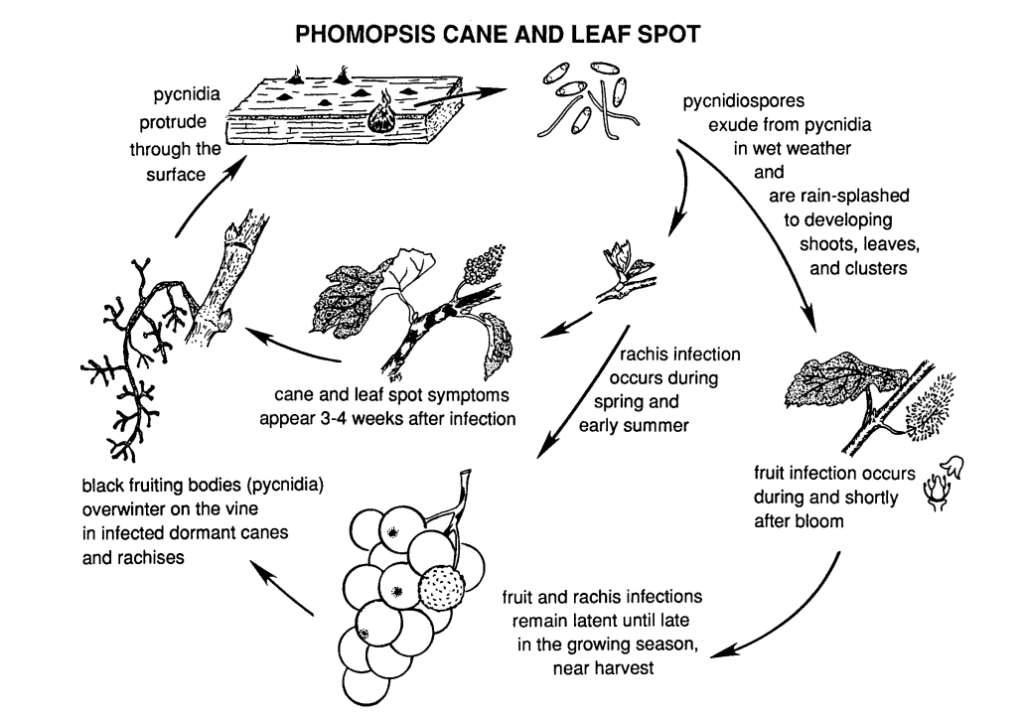
Phomopsisová skvrnitost třtiny a listů révy vinné je způsobována Phomopsis viticola.
Houba přezimuje ve zdřevnatělých částech révy a na jaře následujícího roku je opět aktivní. Jakmile se zvýší teploty a začnou jarní deště, uvolní se spory z přezimujících struktur zvaných pyknidy. Výtrusy se deštěm a větrem rozptylují na nová pletiva a infekce se šíří.
Období primární infekce se obvykle shoduje s časnými jarními dešti a začíná krátce po rašení pupenů, kdy výhony vykazují počáteční fázi růstu. Mladé výhony, vrcholky výhonů, plodenství a obrosty jsou náchylné k infekci. Ačkoli se špičky výhonů mohou infikovat během celé vegetační sezóny, infekce se nejčastěji vyskytují v období od rašení poupat do květu. V plodech může patogen zůstat v klidovém stavu až do dozrání plodů. Inkubační doba obvykle trvá asi 3 až 4 týdny.
Příznaky
Na listech a řapících se objevují malé tmavé skvrny s černým středem obklopené žlutými okraji, které se nakonec spojí. Bazální listy mohou být deformované a pomačkané a nemusí se vyvinout do plné velikosti. Pokud jsou řapíky silně napadeny, žloutnou a opadávají, což vede k opadu listů. Listy vytvářejí deštníkový efekt, který omezuje další infekci sporami.
Podobné skvrny se objevují i na výhonech, kde v místech infekce dochází k praskání epidermálních vrstev. Jak se skvrny spojují, na infikovaných místech výhonů se vytváří strupovitá struktura. V závažných případech mohou výhony zakrnět, zlomit se nebo dokonce odumřít.
Léze na řapíku způsobují jeho propadnutí a křehkost. Po období klidu v létě se houba na začátku podzimu znovu aktivuje a způsobuje hnilobu bobulí a hroznů. Ačkoli infekce plodů nejsou obecně rozsáhlé, na bobulích se tvoří skvrny, které jsou často doprovázeny černými skvrnami (pyknidiemi) na slupce bobulí. Za vlhka mohou z pyknid vytékat žluté výtrusy, což vede ke změně barvy plodů, jejich hrubé struktuře a mumifikaci.
Napadené dřevo se v období vegetačního klidu projevuje vyběleným vzhledem. Silně napadené lodyhy a ostruhy vykazují tmavé zbarvení smíšené s vybělenými plochami v kůře. Pycinidie mohou také prorazit povrch. Nízké teploty spolu s narušenou funkcí floému a xylému oslabují a ničí lodyhy, ostruhy a pupeny.
FieldClimate Model
Vinařství Model infekce Phomopsis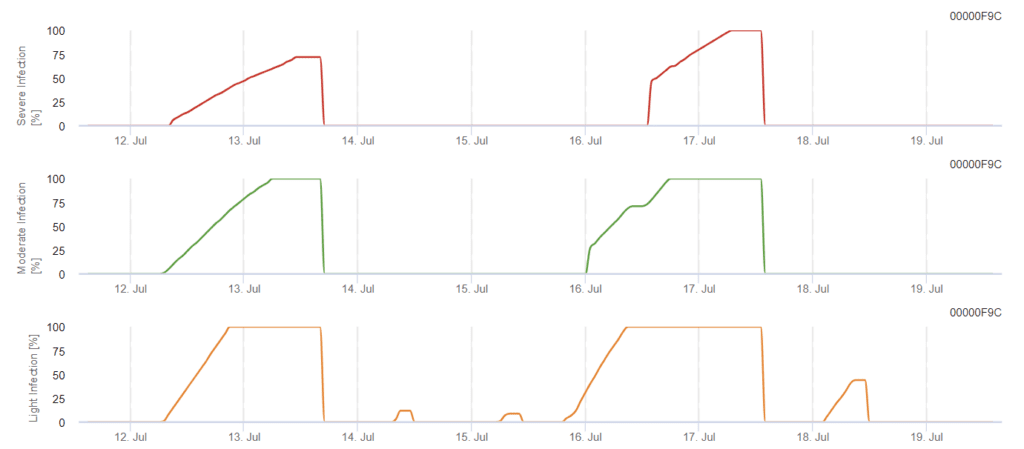
Potřebné senzory:
- Teplota vzduchu
- Relativní vlhkost
- Vlhkost listů
- Srážky
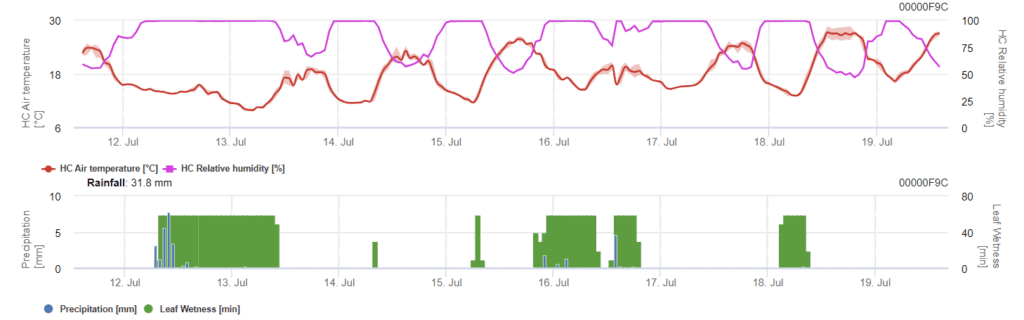
Model určuje riziko infekce Phomopsis při teplotách v rozmezí 5 až 35 °C a vlhkých podmínkách (vlhkost listů, vysoká relativní vlhkost). Závažnost infekce závisí na množství deště (více než 2 mm), protože spory se rychleji šíří do zdravého rostlinného materiálu. Pokud je prokázána infekce 100%, znamená to, že na poli byly naměřeny optimální podmínky pro infekci rostlinného pletiva, a proto je třeba vzít v úvahu měření ochrany rostlin.
Literatura
- https://agriculture.vic.gov.au/biosecurity/plant-diseases/grapevine-diseases/phomopsis-cane-and-leaf-spot-of-grapevines
- Bettiga, L. J. (Ed.). (2013). Ochrana proti škůdcům révy vinné (Vol. 3343). Publikace UCANR.
- Pscheidt, J. W., & Pearson, R. C. (1991). Phomopsis cane & leaf spot.