Práce vychází z následujících dvou článků:
- Choroba černých listových pruhů; Pests and Diseases of American Samoa Number 10, American Samoa Community College Community & Natural Resources Cooperative Research & Extension. 2004
- Černá Sigatoka Stále častěji se vyskytující vlákno pro pěstování banánů; Douglas H. Marín, Del Monte Fresh Produce; Ronald A. Romero, Chiquita Brands; Mauricio Guzmán, National Banana Corporation of Costa Rica; Turner B. Sutton, North Carolina State University, Raleigh
Černá listová skvrnitost (BLS) neboli "černá sigatoka" je celosvětově nejškodlivější choroba listů banánů. Postižena je většina oblastí tropů a subtropů, přičemž jsou hlášeny ztráty ve výši 30-50%. První zpráva o výskytu BLS v Tichomoří pochází z údolí Sigatoka na Fidži, a to před 40 lety. Většina fungicidů používaných v produkci banánů je určena k regulaci BLS. Banán Cavendishova typu "Williams", který pěstuje většina komerčních pěstitelů, je k této houbové chorobě velmi náchylný. Napadené rostliny mají méně listů, což vede k menšímu počtu a menším plodům, opožděné sklizni, předčasnému dozrávání a nižší kvalitě plodů.
Potenciální dopad
Černá sigatoka způsobuje destrukci tkáně banánových listů, což ovlivňuje fotosyntetické schopnosti rostliny a může snížit výnosy až o 50%. Černá sigatoka je jedním z hlavních faktorů způsobujících pokles vývozu banánů v zemích jižního Pacifiku. Komerční plantáže produkující banány na vývoz musí udržovat nákladný program fungicidních postřiků k potlačení černé sigatoky a jsou kritizovány z důvodů ochrany životního prostředí a lidského zdraví. Pokud se však nekontroluje, plody vyprodukované na nemocných rostlinách mohou během přepravy předčasně dozrát a způsobit další ztráty. V následujících letech se podařilo ničivou chorobu potlačit chemickými postřiky (15-17 aplikací fungicidů), což však značně zvýšilo náklady na produkci.
Zasažené rostliny
Definitivní příznaky černé sigatoky byly zaznamenány pouze na Musa (Banány), Musa paradisiaca (Plantejny), Musa acuminata (divoký banán) a Musa acuminata (subsp. bantesii a subsp. zebrina). Odrůdy se liší svou reakcí na patogen. Imunita není známa a je možné, že další volně žijící druhy a poddruhy Musa jsou infikovány, ale onemocnění se výrazně nerozvíjí. Mycosphaerella fijiensis může napadat i sazenice planě rostoucích banánů.
Sezóna výskytu
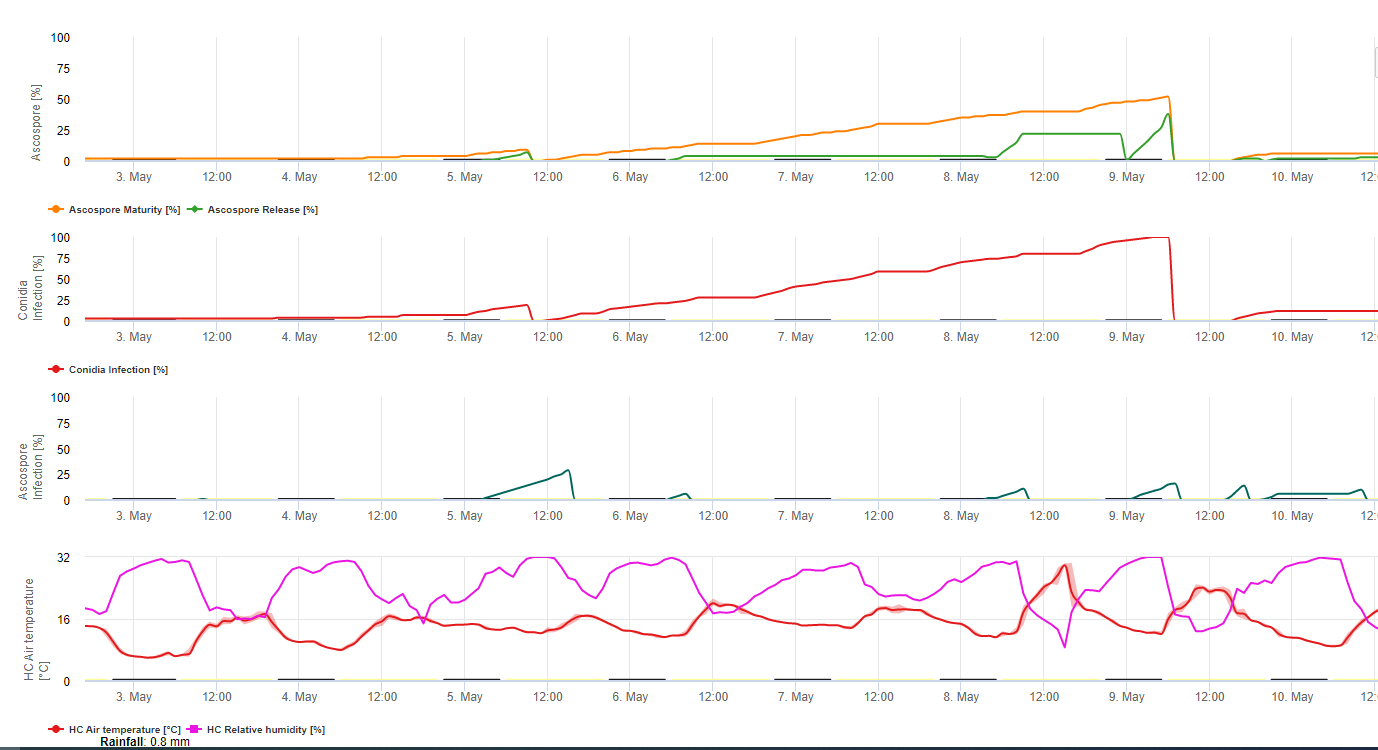
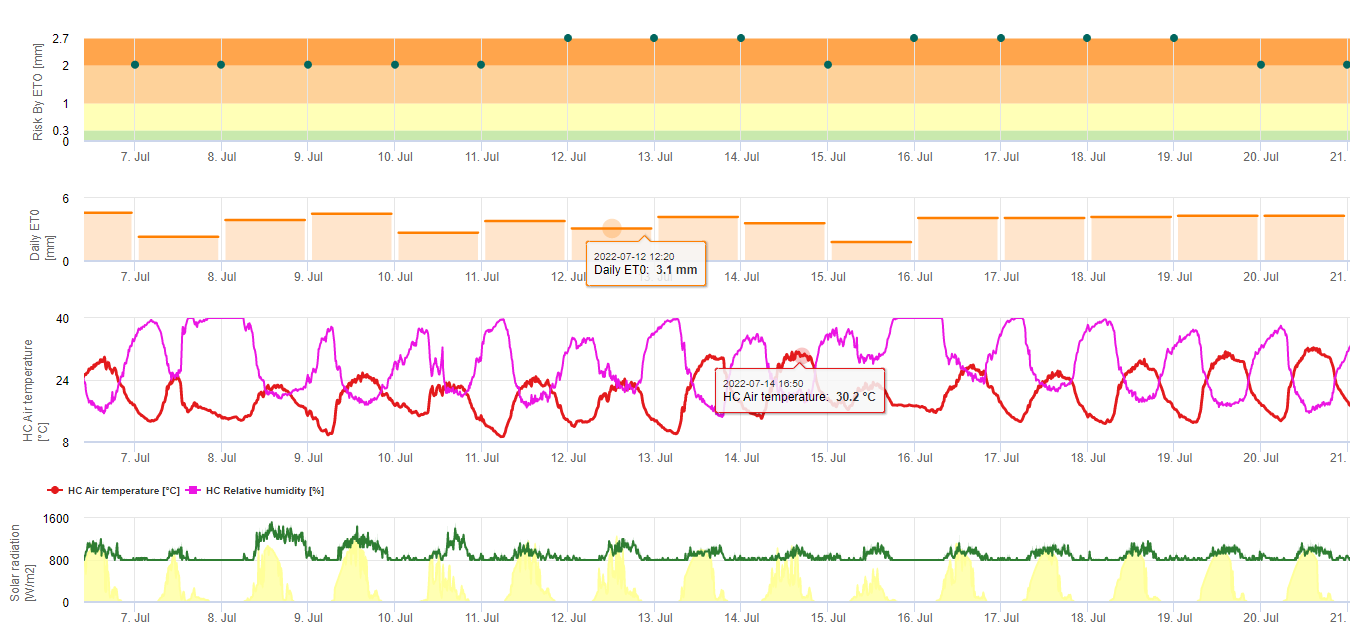
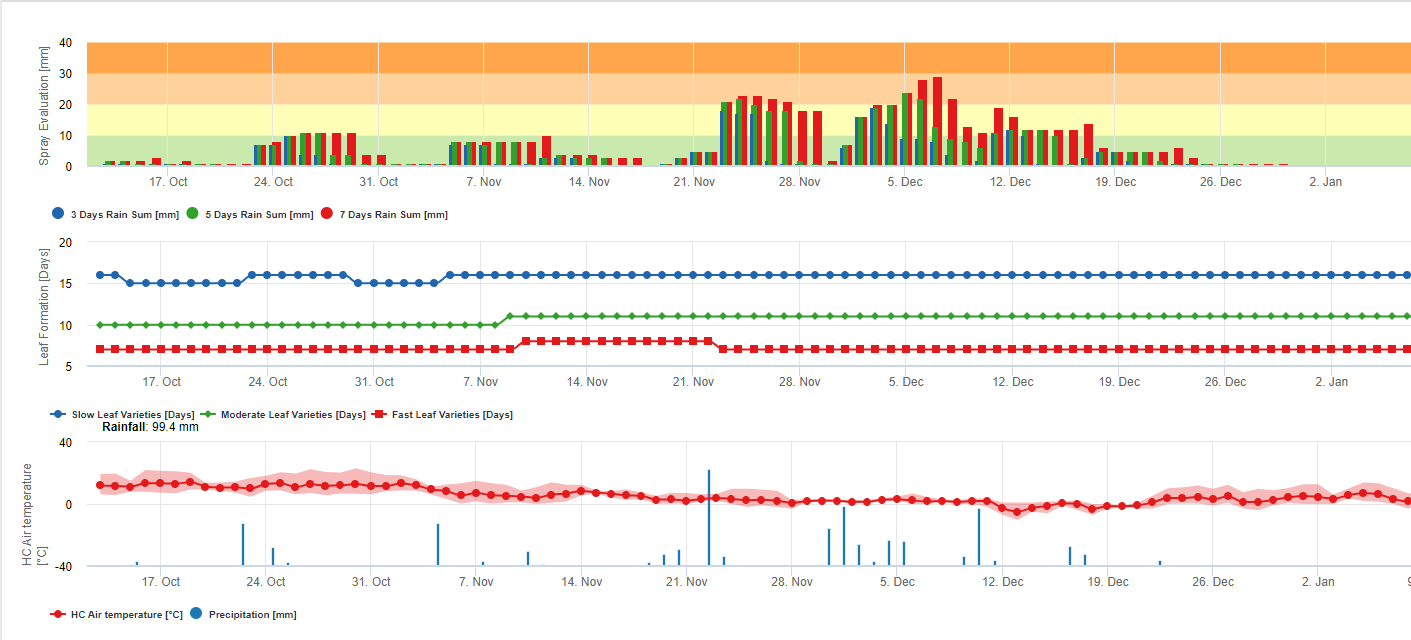
Spory sigatoky černé se snadno tvoří během tropického a subtropického léta nebo v podmínkách vysoká vlhkost a srážky, zejména pokud je na listech volný vodní film. Hlavním způsobem šíření je dešťová voda nebo rozprašování spor, ale v pozdější fázi vývoje choroby jsou spory také násilně uvolňovány do vzdušných proudů. K infekci dochází na nejmladších listech rostliny během rašení a bezprostředně po něm, ale starší listy jsou pro rozvoj choroby příznivější. Zjevné vizuální příznaky se objevují asi 15 až 20 dní po první infekci.
Příznaky
Prvními zjevnými příznaky infekce listů jsou krátké, červenohnědé čáry dlouhé asi 2 mm. Čáry se mění v hnědé pruhy dlouhé 20-30 mm, které se rozšiřují v eliptické skvrny, na spodní straně listu hnědé, na horní straně černé. Skvrny jsou mírně vpadlé a obklopené nažloutlou aureolou. Dále střed velké skvrny zešedne, je ohraničen černou linkou a žlutým svatozáří. Nakonec skvrny splynou a list zahyne.
Epidemiologie
Konidie i askospory se podílejí na šíření choroby. Konidie se tvoří za podmínek vysoké vlhkosti, zejména pokud je na listech volný vodní film. Vytvářejí se během vývoje prvních stadií choroby (zejména během stadia pomlčky 2, stadia pruhu3 a stadia skvrnitosti 4). Hlavními způsoby šíření jsou dešťové smyvy a postřik; konidie se větrem neoddělují. Konidie jsou spojeny především s lokálním šířením choroby a jsou důležité v obdobích vysoké vlhkosti, častých silných ros a občasných přeháněk.
Protože M. fijiensis produkuje relativně málo konidií, jsou askospory považovány za důležitější pro šíření černé Sigatoky (4,20). Jacome et al. však uvádí (10), že konidie jsou schopny způsobit značné množství onemocnění a příznaky onemocnění jsou totožné s příznaky způsobenými infekcí askosporami. Uvedl, že konidie nabývají na významu v suchých obdobích, kdy je vývoj choroby opožděn z důvodu přítomnosti méně příznivých klimatických podmínek (10). Askospory jsou primárním prostředkem rozptyl na větší vzdálenosti v rámci plantáží a do nových oblastí a jsou obvyklým způsobem šíření během delších období vlhkého počasí (3,4,6,10, 11,15).
Askospory se vytvářejí v pseudoteciích ve zralých lézích, které jsou běžné na starších listech rostliny nebo v odumřelých listech ležících na zemi. Meredith a Lawrence (14) uvádějí, že pseudothecia jsou produkována na obou stranách povrchu listů, ale větší počet je přítomen na adaxiálním povrchu. Naopak Gauhl et al. (6) zjistili, že více pseudothecií a askospor se tvoří na abaxiálním povrchu listů. Ačkoli Burt et al. zjistili (2), že na jedno pseudothecium se uvolňuje přibližně 4,5 askospor, zdá se tento počet velmi nízký vzhledem k vysoké koncentraci inokula, které je přítomno ve vzduchu po dešti (4).
Uvolňování askospor
Vyžaduje přítomnost vodní povlak z deště nebo rosy která proniká do pseudotecií a vede k násilnému vyvržení askospor přes mezní vrstvu listů, kde se šíří vzdušnými proudy (15,20). Dozrávání pseudothecií vyžaduje nasycení odumřelých listových tkání po dobu přibližně 48 hodin (3,4,7,10,11,15). V havajských podmínkách se koncentrace askospor zvyšovala během noci, byla nejvyšší kolem 06:00 h a během dne výrazně klesala. V deštivých dnech se nejvyšší koncentrace objevily krátce po začátku deště. Sezónní nárůst průměrných denních koncentrací askospor souvisí se zvýšeným množstvím srážek a relativní vlhkostí vzduchu (15). Gauhl (4), který pracoval s banány, zaznamenal podobné výsledky v kostarických podmínkách. Zatímco Meredith et al. (15) nenašli důkaz, že by produkce nebo uvolňování askospor byly ovlivněny minimálními teplotami, Gauhl (4) zjistil, že v karibské zóně Kostariky dochází ke snížení produkce inokula během sušších (nebo méně deštivých) měsíců roku, což je zároveň období s nejnižšími teplotami. Askospory se šíří větrem, avšak jejich šíření na velké vzdálenosti je omezeno na několik set kilometrů kvůli jejich citlivosti na ultrafialové záření (17). Nebyla prokázána konzistentní souvislost mezi rozptylem askospor a rozvojem choroby, což omezuje užitečnost odchytu spor pro předpověď choroby (5,11). Askospory se během rozvíjení nového listu ukládají hlavně na spodním povrchu listu a vytvářejí pásovou strukturu infekcí na straně, která je odhalena jako první, což je odrazem zvýšeného ukládání spor na válcovitém listu kandíku během jeho rozvíjení na rozdíl od celé otevřené listové desky. Většina infekcí se proto vyskytuje na abaxiálním povrchu listů (3,7).
Infekce
Zárodečné trubice pronikají do průduchů přibližně za 48 až 72 hodin (1,20), ačkoli houba může růst epifyticky na povrchu listů až 6 dní, než skutečně pronikne do listů (1). Úspěšná infekce je podporován rozšířenou období vysoké vlhkosti a přítomnosti volné vody na listech. (3,7,13). K maximální klíčivosti dochází při přítomnosti volné vody. Konidie klíčí v širším rozmezí relativní vlhkosti (92 až 100%) než askospory (98 až 100%). Vliv teplota na klíčení lze charakterizovat kvadratickou funkcí odezvy s odhadovaným optimem 26,5 °C (10). Stover (21) pozoroval maximální růst zárodečných trubic askospor při teplotě 26 až 28 °C po 24hodinové inkubaci. Jacome a Schuh (8,9) uvádějí, že starší listy jsou náchylnější; E. Bureau (osobní sdělení) a Romero (18) však pozorovali, že mladší listy jsou náchylnější (první až třetí) než starší.
Inkubační doba
Doba mezi infekcí a objevením se příznaků se liší v závislosti na povětrnostních podmínkách a citlivosti rostlin (68). U banánů se doba infekce listů odhaduje na dobu, kdy se objeví nový list z vrcholu pseudostomu (20). Za velmi příznivých podmínek v Kostarice a u vnímavého hostitele se může vyskytnout inkubační doba může trvat jen 13 až 14 dní, zatímco v období nepříznivého počasí se může inkubační doba prodloužit až na 35 dní. Podobné zprávy existují i z Nigérie o plantejnech (16). V období dešťů byla inkubační doba 14 dní, ale v období sucha 24 dní. Délka inkubační doby se také liší v závislosti na citlivosti odrůdy k M. fijiensis. U kultivarů FHIA 1 a FHIA 2, dvou tetraploidních syntetických hybridů vyvinutých FHIA, byla inkubační doba 26,2 a 25,1 dne, zatímco u náchylného kultivaru Grande Naine byla 22,4 dne (19).
Po proniknutí do listu se hyfy <M. fijiensis kolonizovat sousední buňky po dobu přibližně 7 dnů bez jakýchkoli známek narušení buněk. Vegetativní hyfy mohou vyrůstat ze stomatu a růst na povrchu listu a pronikat do sousedních stomat nebo vytvářet konidiofory a konidie. Tento epifytický růst umožňuje houbě kolonizovat přilehlé listové pletivo, což vede k rychlému rozvoji symptomů (4,20). M. fijiensis má větší schopnost proniknout několika žaludky, než má M. musicola, což má za následek větší skvrnitost než u prvního patogenu (20).
Latentní období
Ačkoli konidie mohou být produkovány v lézích vykazujících časné příznaky M. fijiensis a přispívají k epidemii, je latentní období vymezeno dobou, kdy houba začne vytvářet léze se zralými pseudotécii a askosporami, které jsou hlavním zdrojem inokula. Stejně jako inkubační doba se i latentní doba mění v závislosti na povětrnostních podmínkách, vnímavosti hostitele a intenzitě infekce. Latentní doba se pohybovala od 25 dnů v období dešťů (červen až prosinec) do 70 dnů v období sucha v Guapiles na Kostarice. Když je počasí velmi příznivé pro vylučování askospor a infekci, dochází k mnoha infekcím na listech. Když jsou infekce husté, rychle se spojují ve velmi raném stádiu vývoje, což urychluje vznik zralých skvrn, které se vyznačují přítomností pseudothecií a askospor (22). Za těchto podmínek jsou listy rychle a vážně poškozeny. Latentní období se rovněž liší v závislosti na úrovni odolnosti. Například doba od objevení se listů do prvního příznaku zralé skvrnitosti byla za stejných přírodních podmínek u kultivaru Curraré, banánu na vaření patřícího do podskupiny plantain, 44 dní ve srovnání s 34 dny u kultivaru Valery, banánu patřícího do podskupiny Cavendish (4). Termín "doba vývoje choroby" se v literatuře o banánech v tropech běžně používá pro latentní období a je definován jako doba mezi infekcí a vznikem zralých skvrn. Dalším termínem běžně používaným v literatuře o banánech je "doba vývoje příznaků" nebo "přechodné období", což je doba od prvních příznaků do vzniku zralých skvrn (6). Doba vývoje příznaků dobře ukazuje, jak rychle choroba na listech postupuje.
Literatura:
- Beveraggi, A. 1992. Etude des interactions hôte-parasite chez des bananiers sensibles et résistants inoculés par Cercospora fijiensis responsible de la maladie des raies noires. Thèse de 3ème cycle. Université de Montpellier, Montpellier, Francie.
- Burt, J. P. A., Rosenberg, L. J., Rutter, J., Ramírez, F. a González, O. H. 1999. Forecasting the airborne spread of Mycosphaerella fijiensis, cause of black Sigatoka disease on banana (Předpověď šíření Mycosphaerella fijiensis, původce černé sigatoky na banánech, vzduchem): Odhady počtu perithecií a askospor. Ann. Appl. Biol. 135:369-377.
- Fullerton, R. A. 1994. Sigatoka Leaf Diseases.Pages 12-14 in: B.: Compendium of Tropical Fruit Diseases. R. C. Ploetz, G. A. Zentmyer, W. T. Nishijinia, K. G. Rohrbach, and H. D. Ohr, eds. American Phytopathological Society, St. Paul, MN.
- Gauhl, F. 1994. Epidemiology and Ecology of black Sigatoka (Mycosphaerella fijiensis Morelet) on Plantain and Banana (Musa spp.) in Costa Rica, Central America. INIBAP, Montpellier, Francie.
- Gauhl, F. 2002. Jsou lapače spor užitečným nástrojem pro předpověď vývoje choroby Sigatoka na komerčních banánových plantážích. Strana 22 v: Int. Workshop Mycosphaerella Leaf Spot Dis. Bananas, 2nd - Programme and Abstracts. (Abstr.)
- Gauhl, F., Pasberg-Gauhl, C. a Jones, D. R. 2000. Cyklus onemocnění a epidemiologie. Pages 56-62 in: Diseases of Banana, Abacá and Enset. D. R. Jones, ed. CAB International, Wallingford, Velká Británie.
- González, M. 1987. Enfermedades del cultivo del banano. Oficina de Publicaciones de la Universidad de Costa Rica, San José.
- Jacome, L. H. a Schuh, W. 1992. Vliv délky trvání vlhkosti listů a teploty na vývoj choroby černá Sigatoka na banánech napadených Mycosphaerella fijiensis var. difformis. Phytopathology 82:515-520.
- Jacome, L. H. a Schuh, W. 1993. Vliv teploty na růst a produkci konidií in vitro a srovnání infekce a agresivity in vivo mezi izoláty Mycosphaerella fijiensis var. difformis. Trop. Agric. 70:51-59.
- Jacome, L. H., Schuh, W. a Stevenson, R. E. 1991. Vliv teploty a relativní vlhkosti na klíčení a vývoj zárodečných trubic Mycosphaerella fijiensis var. difformis. Phytopathology 81:1480-1485.
- Jeger, M. J., Eden-Green, S., Thresh, J. M., Johanson, J. M., Waller, J. M. a Brown, A. E. 1995. Banana diseases. Pages 317-381 in: Bananas and Plantains. S. Gowen, ed. Chapman & Hall, London.
- Marín, D. H., a Romero, R. A. 1992. El combate de la Sigatoka negra. Boletín No. 4, Departamento de Investigaciones, Corporación Bananera Nacional, Costa Rica.
- Mayorga, M. H. 1990. La Raya negra (Mycosphaerella fijiensis Morelet) del plátano y del banano. I. Ciclo de vida del patógeno bajo las condiciones del Urabá. Revista ICA 25:69-77.
- Meredith, D. S., a Lawrence, J. S. 1969. Black leaf streak disease of bananas (Mycosphaerella fijiensis): Symptoms of disease in Hawaii, and notes on the conidial state of the causal fungus. Trans. Br. Mycol. Soc. 52:459-476.
- Meredith, D. S., Lawrence, J. S., a Firman,I. D. 1973. Uvolňování a šíření askospor u černé listové skvrnitosti banánů (Mycosphaerella fijiensis). Trans. Br. Mycol. Soc. 60:547-554.
- Mobambo, K. N., Gauhl, F., Pasberg-Gauhl, C. a Zuofa, K. 1996. Hodnocení vlivu sezóny a věku rostlin u planě rostoucího jitrocele na reakci na chorobu černá Sigatoka. Crop Prot. 15:609- 614.
- Parnell, M., Burt, P. J. A. a Wilson, K. 1998. The influence of exposure to ultraviolet radiation in simulated sunlight on ascospores causing black Sigatoka disease of banana and plantain (Vliv vystavení ultrafialovému záření v simulovaném slunečním světle na askospory způsobující černé onemocnění banánů a plantainů). Int. J. Biometeorol. 42:22-27.
- Romero, R. A. 1995. Dynamika populací Mycosphaerella fijiensis odolných vůči fungicidům a epidemiologie černé sigatoky banánů. Ph.D. disertační práce. North Carolina State, Raleigh.
- Romero, R. A. a Sutton, T. B. 1997. Reakce čtyř genotypů Musa při třech teplotách na izoláty Mycosphaerella fijiensis z různých oblastí. Plant Dis. 81:1139-1142.
- Stover, R. H. 1980. Sigatoka leaf spot of bananas and plantains (Sigatoková skvrnitost listů banánů a plantejnů). Plant Dis. 64:750-756.
- Stover, R. H. 1983. The effect of temperature on ascospore germ tube growth of Mycosphaerella musicola and Mycosphaerela fijiensis var. difformis. Fruits 38:625-628.