

O míldio da Batata e do Tomate
Randall C. Rowe, Sally A. Miller, Richard M. Riedel, Ohio State University Extension Service
O míldio é uma doença muito comum tanto da batata como do tomate. Causa manchas foliares e manchas de tubérculo na batata, e manchas foliares, podridão dos frutos e lesões no caule do tomate. A doença pode ocorrer numa vasta gama de condições climáticas e pode ser muito destrutiva se não for controlada, resultando muitas vezes numa desfoliação completa das plantas. Em contraste com o nome, raramente se desenvolve cedo, mas geralmente aparece na folhagem madura.
Sintomas
Nas folhas de ambas as culturas, os primeiros sintomas aparecem geralmente nas folhas mais velhas e consistem em pontos mortos pequenos, irregulares, castanhos escuros a pretos, de tamanho que varia entre um ponto preciso e 1/2 polegada de diâmetro. À medida que as manchas se alargam, podem formar-se anéis concêntricos como resultado de padrões de crescimento irregulares pelo organismo no tecido da folha. Isto dá à lesão um aspecto característico de "ponto alvo" ou "olho de touro". Há frequentemente uma auréola estreita e amarela à volta de cada mancha e as lesões são normalmente delimitadas por veias. Quando as manchas são numerosas, podem crescer juntas, fazendo com que as folhas infectadas fiquem amarelas e morram. Normalmente, as folhas mais antigas tornam-se infectadas primeiro e secam e caem da planta à medida que a doença progride pelo caule principal.
No tomate, as infecções do caule podem ocorrer em qualquer idade, resultando em áreas pequenas, escuras, ligeiramente afundadas, que se alargam para formar pontos circulares ou alongados com centros de cor mais clara. Marcas concêntricas, semelhantes às das folhas, desenvolvem-se frequentemente em lesões do caule. Se forem utilizadas sementes infestadas para iniciar transplantes de tomate, as plântulas podem ficar húmidas logo após a sua emergência. Quando grandes lesões se desenvolvem na linha do solo em caules de transplantes ou plântulas, as plantas podem ficar cingidas, uma condição conhecida como "podridão do colar". Tais plantas podem morrer quando colocadas no campo ou, se os caules estiverem enfraquecidos, podem partir-se no início da estação. Algumas plantas podem sobreviver com sistemas radiculares reduzidos se porções de caules acima do cancro desenvolverem raízes onde entram em contacto com o solo. Tais plantas, contudo, produzem geralmente poucos ou nenhuns frutos. As lesões nos caules são muito menos comuns e destrutivas na batata.
A queda da flor e a mancha de caules de frutos, juntamente com a perda de frutos jovens, podem ocorrer quando a floração precoce ataca os tomates na fase de floração. Nos frutos mais velhos, o míldio provoca manchas escuras e de pele afundada, geralmente no ponto de fixação do caule. Estas manchas podem aumentar para envolver toda a parte superior do fruto, mostrando frequentemente marcas concêntricas como as das folhas. As áreas afectadas podem ser cobertas com massas negras aveludadas de esporos. Os frutos podem também ser infectados na fase verde ou madura através de fissuras de crescimento e outras feridas. Os frutos infectados caem frequentemente antes de atingirem a maturidade.
Nos tubérculos de batata, o míldio da erva resulta em lesões superficiais que parecem um pouco mais escuras do que a pele saudável adjacente. As lesões são geralmente ligeiramente afundadas, circulares ou irregulares, e variam em tamanho até 3/4 polegadas de diâmetro. Existe normalmente uma margem bem definida e por vezes ligeiramente elevada entre tecido saudável e doente. Internamente, o tecido apresenta uma podridão castanha a negra, seca, geralmente não mais de 1/4 a 3/8 polegadas de profundidade. Podem formar-se fissuras profundas em lesões mais antigas. A infecção do tubérculo é invulgar em condições de Ohio.
Pathogen
A praga precoce é causada pelo fungo, Alternaria solani, que sobrevive em folhas ou tecidos de caule infectados no solo ou no solo. Este fungo está universalmente presente nos campos onde estas culturas têm sido cultivadas. Também pode ser transportado em sementes de tomate e em tubérculos de batata. Formam-se esporos em detritos vegetais infestados à superfície do solo ou em lesões activas numa gama de temperaturas bastante ampla, especialmente em condições alternadas de humidade e seca. São facilmente transportados por correntes de ar, solo soprado pelo vento, chuva salpicada, e água de irrigação. A infecção de folhas ou tecidos de caule susceptíveis ocorre em tempo quente e húmido com orvalho ou chuva intensa. O míldio pode desenvolver-se muito rapidamente em meados ou finais da estação e é mais grave quando as plantas são stressadas por má nutrição, seca, ou outras pragas. A infecção dos tubérculos de batata ocorre através de aberturas naturais na pele ou através de lesões. Os tubérculos podem entrar em contacto com esporos durante a colheita e as lesões podem continuar a desenvolver-se no armazenamento.
TOMCAST (Jim Jasinski, Coordenador do TOMCAST para OHIO, INDIANA, & MICHIGAN)
Antecedentes
TOMCAST (TOMato disease foreCASTing) é um modelo informático baseado em dados de campo que tenta prever o desenvolvimento de doenças fúngicas, nomeadamente o Early Blight, Septoria Leaf Spot e Anthracnose no tomate. Os registadores de dados colocados no campo estão a registar dados horários de humidade das folhas e temperatura. Estes dados foram analisados durante um período de 24 horas e podem resultar na formação de um valor de severidade da doença (DSV); essencialmente um incremento do desenvolvimento da doença. À medida que o DSV se acumula, a pressão da doença continua a aumentar sobre a cultura. Quando o número de DSV acumulado excede o intervalo de pulverização, recomenda-se uma aplicação de fungicida para aliviar a pressão da doença.
TOMCAST
Aplicações de fungicidas de cronometragem para o flagelo precoce, Septoria leaf spot, e Anthracnose
Um sistema de previsão de doenças baseado no tempo chamado TOMCAST, desenvolvido pelo Dr. Ron Pitblado no Ridgetown College of Agricultural Technology em Ontário, Canadá, pode ser usado para cronometrar aplicações fungicidas para três doenças fúngicas: a ferrugem precoce (causada por Alternaria solani), a mancha foliar Septoria (causada por Septoria lycopersici), e a antracnose da fruta (causada por Colletotrichum coccodes). Se o late blight estiver presente no seu condado ou condados adjacentes, ou se estiverem presentes condições para o movimento de esporos na sua área, utilize o sistema de previsão do Simcast late blight para cronometrar as aplicações fungicidas.
NOTA CAUTELAR IMPORTANTE:
O TOMCAST não é útil em quintas que têm um historial de doenças bacterianas. Se normalmente tem problemas com manchas bacterianas, manchas, ou cancro nos seus tomates, não deve usar TOMCAST porque os intervalos de pulverização recomendados não serão suficientes para o controlo de doenças bacterianas se estiver a misturar o cobre com as suas aplicações fungicidas. Encontre aqui informações sobre como reduzir as doenças bacterianas: http://extension.psu.edu/plants/vegetable-fruit/news/2015/farming-like-you-expect-bacterial-diseases
TOMCAST utiliza dados de humidade e temperatura das folhas para calcular os valores de gravidade da doença (DSV's), como se mostra no Quadro 1.
TOMCAST é derivado do modelo original F.A.S.T. (Forecasting Alternaria solani on Tomatoes) desenvolvido por Drs. Madden, Pennypacker, e MacNab ? na Universidade Estadual da Pensilvânia (PSU). O modelo PSU F.A.S.T. foi ainda modificado pelo Dr. Pitblado no Ridgetown College em Ontário, no que agora reconhecemos como o modelo TOMCAST utilizado pela Ohio State University Extension.
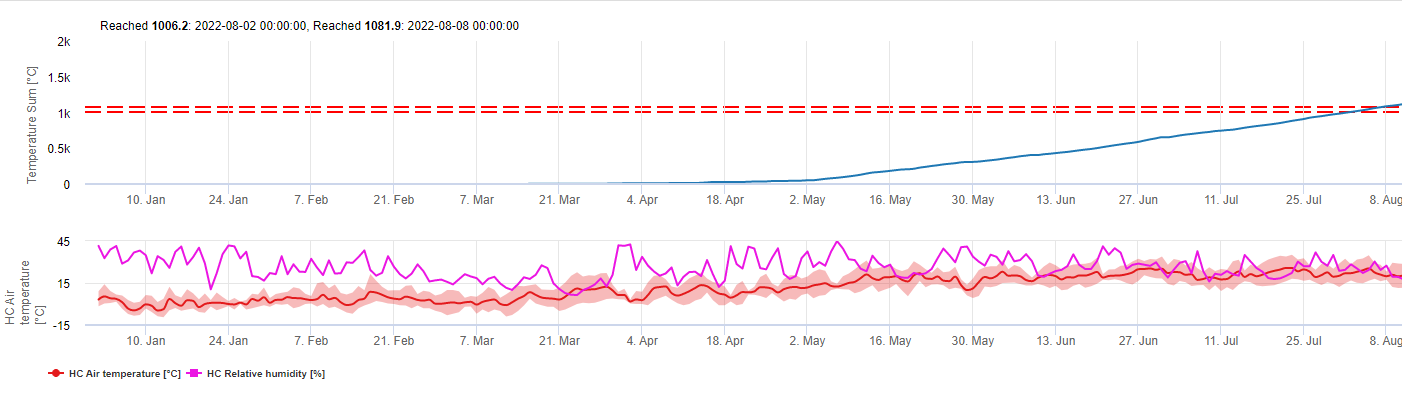
DSV são A Disease Severity Value (DSV) é a unidade de medida dada a um incremento específico do desenvolvimento da doença (Early Blight). Por outras palavras, um DSV é uma representação numérica do quão rápido ou lento a doença (Early Blight) se está a acumular num campo de tomate. O DSV é determinado por dois factores; humidade das folhas e temperatura durante as horas "leaf wet". À medida que o número de horas de "folha molhada" e a temperatura aumentam, o DSV acumula a um ritmo mais rápido. Ver a tabela de valores de severidade da doença abaixo.
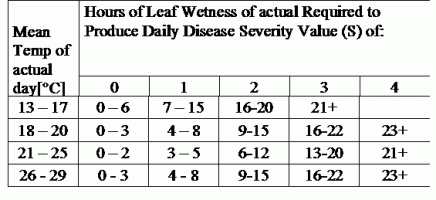
Inversamente, quando há menos horas húmidas de folhas e a temperatura é mais baixa, o DSV acumula-se lentamente, se é que se acumula. Quando o número total de DSV acumulados excede um limite predefinido, chamado intervalo ou limiar de pulverização, recomenda-se um spray fungicida para proteger a folhagem e a fruta do desenvolvimento de doenças.

O intervalo de pulverização (que determina quando se deve pulverizar) pode variar entre 15-20 DSV. O DSV exacto que um produtor deve utilizar é normalmente fornecido pelo processador e depende da qualidade da fruta e da utilização final dos tomates. Seguindo um intervalo de 15 DSV spray é uma utilização conservadora do sistema TOMCAST, o que significa que pulverizará mais frequentemente do que um cultivador que utiliza um intervalo de 19 DSV spray com o sistema TOMCAST. A troca está no número de pulverizações aplicadas durante a estação e no potencial de diferença na qualidade da fruta.
UTILIZAÇÃO DO TOMCAST
Os tomates cultivados num raio de 10 milhas de uma estação de notificação devem beneficiar da função de gestão de doenças do TOMCAST para ajudar a prever o flagelo precoce, Septoria, e Antracnose. Se decidir experimentar TOMCAST nesta estação, por favor tenha em mente três conceitos muito importantes.
Um: Se esta é a primeira vez que utiliza o sistema, recomenda-se que apenas parte da sua área seja colocada no programa para ver como se ajusta aos seus padrões de qualidade e estilo operacional.
Dois: Use TOMCAST como guia para ajudar a melhorar as aplicações fungicidas de tempo, percebendo que em algumas estações pode realmente aplicar mais produto do que um programa de horário definido pode exigir.
Três: Quanto mais um campo de tomate provém de um local de notificação aumenta a probabilidade de distorção na acumulação de DSV, ou seja, o valor notificado pode ser alguns DSV mais alto ou mais baixo do que o verificado pela localização do campo. Isto deve ser tomado em consideração quando a aplicação de fungicidas está provavelmente a alguns dias de distância. Ouça os relatórios DSV das estações próximas e triangule para o seu próprio local como a melhor forma de estimar aproximadamente a sua acumulação de DSV.
PRIMEIRA PULVERIZAÇÃO USANDO TOMCAST
Tem havido alguma discussão ao longo dos anos sobre a aplicação do primeiro spray quando se seguiu o TOMCAST. A regra estabelecida no Guia de Produção Vegetal de 1997 centra-se em torno da data de plantio.
As plantas de tomateiro que entram no campo antes de 20 de Maio devem ter o primeiro spray aplicado quando o DSV para essa área exceder 25 ou quando chegar uma data segura de falha de 15 de Junho. O cofre de segurança só é utilizado se não tiver sido tratado desde 20 de Maio, e é um meio de eliminar o inóculo inicial da doença. Após a primeira pulverização, estes tomates são subsequentemente tratados quando o intervalo de pulverização escolhido (intervalo 15-20 DSV) é excedido.
Os tomates plantados após 20 de Maio são tratados quando excedem o intervalo de pulverização escolhido (intervalo 15-20 DSV) ou quando não foram tratados até à data de segurança de 15 de Junho. Por conseguinte, é fundamental comparar a data de plantação do tomate com a data de início do relatório DSV nessa área para orientar o processo de decisão de pulverização.
Há duas grandes doenças de Septoria no trigo. Estas são a mancha Septoria tritici, incitada pelo fungo Septoria tritici (teleomorfo: Mycophaerella graminicola), e Septoria nodorum mancha, causada pelo fungo Septoria nodorum (teleomorph: Leptosphaeria nodorum). Ambas as doenças causam graves perdas de rendimento relatadas entre 31 e 53 por cento (Eyal, 1981; Babadoost e Herbert, 1984; Polley e Thomas, 1991). A nível mundial, mais de 50 milhões de hectares de trigo, que crescem principalmente nas zonas de grande pluviosidade, são afectados. Durante os últimos 25 anos, estas doenças têm vindo a aumentar e tornaram-se um importante factor limitador da produção de trigo em certas áreas. Sob epidemias graves, os grãos de cultivares de trigo susceptíveis são murchos e não estão aptos para moagem. As epidemias de Septoria tritici blotch e Septoria nodorum blotch de trigo estão associadas a condições climáticas favoráveis (chuvas frequentes e temperaturas moderadas), práticas culturais específicas, disponibilidade de inóculos e presença de cultivares de trigo susceptível (Eyal et al., 1987).
Septoria spp. Biologia
Após Erick De Wolf, Septoria Tritici Blotch, Kansas State University, Abril de 2008 Septoria tritici mancha conhecida como mancha de folha manchada, é causada pelo fungo Septoria tritici. Está distribuído em todas as zonas de cultivo de trigo do mundo e é um problema grave em muitas regiões. A mancha Septoria tritici é mais prejudicial quando a doença ataca as folhas superiores e as cabeças das variedades susceptíveis no final da estação.
Sintomas
Septoria tritici Os sintomas da mancha aparecem pela primeira vez no Outono. Os sintomas iniciais são pequenas manchas amarelas nas folhas. Estas lesões tornam-se frequentemente bronzeadas leves à medida que envelhecem, e os corpos de frutificação fúngica podem ser vistos embutidos nas lesões nos awns. As lesões têm forma irregular e variam de elípticas a longas e estreitas (Figura 1). As lesões contêm pequenas manchas redondas e pretas que são os corpos de frutificação do fungo. Os corpos de frutificação negros parecem grãos de pimenta preta e podem geralmente ser vistos sem a ajuda de uma lupa. A doença começa nas folhas inferiores e progride gradualmente até à folha da bandeira. As bainhas das folhas também são susceptíveis de ataque. Em anos húmidos, o fungo mancha foliar manchado pode deslocar-se para as cabeças e causar lesões castanhas nas glumas e nas mucosas conhecidas como mancha de glume. Estas lesões tornam-se frequentemente bronzeadas leves à medida que envelhecem e os corpos de frutificação fúngica são frequentemente vistos embutidos nas lesões nos awns.
A fase de mancha de calcário pode causar perda significativa de rendimento, mas a relação entre a gravidade da doença e a perda de rendimento não é bem compreendida. Septoria tritici mancha pode ser confundida com outras doenças das folhas que têm sintomas muito semelhantes: mancha de bronzeado e Stagonspora nodorum mancha, por exemplo. É comum que as plantas sejam infectadas por mais do que uma destas doenças foliares, e pode ser necessário um exame laboratorial para diagnosticar com precisão quais as doenças mais prevalecentes. O exame laboratorial é quase sempre necessário para distinguir a causa da mancha de calcário. Conhecer as espécies não é importante para as decisões de pulverização, porque as três doenças respondem de forma semelhante aos fungicidas. No entanto, saber que doenças são mais prevalentes é uma parte importante da selecção de variedades, porque diferentes genes controlam a resistência às doenças.
A forma mais fiável de distinguir a Septoria tritici blotch das outras doenças é através da presença dos corpos de frutificação de fungos negros. O fungo que provoca a mancha de bronzeado não produz este tipo de estrutura reprodutiva. Contudo, em condições húmidas, o fungo que causa a mancha de Stagonospora nodorum produzirá corpos de frutificação castanhos-claros. Para além da diferença de cor, estas estruturas são também menores do que as produzidas pela Septoria tritici.
Ciclo de vida
Septoria tritici sobrevive durante todo o Verão aos resíduos de uma colheita anterior de trigo e inicia infecções no Outono. Há algumas provas de que o fungo é capaz de sobreviver em associação com outros hospedeiros de gramíneas e sementes de trigo. Estas fontes do fungo são provavelmente mais importantes quando os resíduos do trigo estão ausentes. Independentemente da rotação ou das práticas de gestão de resíduos, existe normalmente inóculo suficiente para iniciar infecções na queda. Septoria tritici A mancha é favorecida pelo tempo fresco e húmido. A gama óptima de temperaturas é de 16 a 21 °C; no entanto, podem ocorrer infecções durante os meses de Inverno a temperaturas tão baixas como 5 °C. A infecção requer pelo menos 6 horas de humidade das folhas, e até 48 horas de humidade são necessárias para uma infecção máxima. Uma vez ocorrida a infecção, o fungo leva 21 a 28 dias para desenvolver os corpos de frutificação negra característicos e produzir uma nova geração de esporos. Os esporos produzidos nestes corpos de frutificação são exsudados em massas pegajosas e requerem chuva para os espirrar sobre as folhas superiores e cabeças.
Infecção por Septoria tritici
Pycnidiosporos de S. tritici germinar em água livre de ambas as extremidades do esporo ou de células intercalares (Weber, 1922). A germinação dos esporos só começa cerca de 12 horas após o contacto com a folha. Os tubos germinativos crescem aleatoriamente sobre a superfície da folha. Weber (1922) observou apenas a penetração directa entre as células epidérmicas, mas outros concluíram que a penetração através de estomas abertos e fechados é o principal meio de penetração do hospedeiro (Benedict, 1971; Cohen e Eyal, 1993; Hilu e Bever, 1957). Kema et al. (1996) observaram apenas a penetração estomatológica. As hifas que crescem através dos estomas tornam-se apertadas a cerca de 1 μm de diâmetro, tornando-se depois mais largas após atingirem a cavidade substomatal.
As hifas crescem paralelamente à superfície da folha sob as células epidérmicas, depois através da mesofila para as células da epiderme inferior, mas não para a epiderme. Não se formam haustorias e o crescimento hifálico é limitado pelas células esclerênquimas em redor dos feixes vasculares, excepto quando as hifas são muito densas. Os feixes vasculares não são invadidos. As hifas crescem intercelularmente ao longo das paredes celulares através da mesofila, ramificando-se num septo ou no meio de uma célula. Não aparecem sintomas macroscópicos durante cerca de 9 dias, excepto para uma célula ocasionalmente morta, mas as células mesofílicas morrem rapidamente após 11 dias. As picnídeas desenvolvem-se em câmaras substomatais. As hifas raramente crescem em células hospedeiras (Hilu and Bever, 1957; Kema et al, 1996; Weber, 1922).
A infecção bem sucedida só ocorre após pelo menos 20 horas de elevada humidade. Apenas algumas manchas castanhas se desenvolvem se as folhas permanecerem húmidas durante 5-10 horas após a deposição de esporos (Holmes e Colhoun, 1974) ou até 24 horas (Kema et al., 1996). As relações hospedeiro-parasita são as mesmas em trigo resistente ou susceptível. A germinação de esporos na superfície da folha é a mesma, independentemente da susceptibilidade. O número de penetrações bem sucedidas é aproximadamente o mesmo, mas o crescimento hifálico é mais rápido em cultivares susceptíveis, resultando em mais lesões. As hifas estendem-se 44 Sessão 2 - B.M. Cunfer para além da área necrótica em todas as cultivares. Uma toxina pode desempenhar um papel na patogénese (Cohen e Eyal, 1993; Hilu e Bever, 1957). Em contraste, a colonização foi grandemente reduzida numa linha resistente (Kema et al., 1996).
Stagonospora (Septoria) e Septoria Pathogens de Cereais: O Processo de Infecção
B.M. Cunfer, Departamento de Fitopatologia, Universidade da Geórgia, Griffin, GA
O processo de infecção foi estudada mais intensamente para o Stagonospora (Septoria) nodorum e Septoria tritici. Um estudo aprofundado sobre Septoria passerinii está disponível. Quase toda a informação relatada é relativa à infecção por picnidiosporos. No entanto, o processo de infecção por outras formas de esporos é bastante semelhante. A informação apresentada é sobretudo para a infecção de folhas em condições óptimas. Alguns estudos foram feitos com plantas de plântulas intactas, enquanto outros foram conduzidos com folhas descoladas. Infecção do coleóptero de trigo e das plântulas por S. nodorum foi descrito em pormenor por Baker (1971) e revisto por Cunfer (1983). Embora não tenham sido feitas comparações precisas, parece que o processo de infecção tem muitas semelhanças em cada sistema hospedeiro-parasita e é típico de muitos agentes patogénicos necrotróficos. As informações sobre factores que influenciam o desenvolvimento dos sintomas e a expressão da doença estão excluídas, mas foram revistas por outros autores (Eyal et al., 1987; King et al., 1983; Shipton et al., 1971). Inclui-se um resumo dos factores que afectam a longevidade dos esporos na superfície da folha.
Papel do Cirrus e da Sobrevivência dos Esporos sobre a superfície da folha A informação mais detalhada sobre a função dos cirros que revestem os picnidiosporos exsudados do picnídio é para S. nodorum. O cirrus é um gel composto por compostos proteicos e sacarídeos. A sua composição e função são semelhantes às de outros fungos nos Sphaeropsidales (Fournet, 1969; Fournet et al., 1970; Griffiths e Peverett, 1980). Os principais papéis dos componentes dos cirros são a protecção dos picnidiosporos contra a dessecação e a prevenção da germinação prematura.
O cirrus protege os picnídeos para que alguns permaneçam viáveis pelo menos 28 dias (Fournet, 1969). Quando o cirro foi diluído com água, se a concentração da solução de cirro era >20%, menos que 10% de picnidiosporos germinou. Com uma concentração menor, os componentes fornecem nutrientes que estimulam a germinação dos esporos e o alongamento dos tubos germinativos. O comprimento dos tubos de germes aumentou até 15% de concentração de cirrus, depois diminuiu moderadamente em concentrações mais elevadas (Harrower, 1976). Brennan et al. (1986) relataram uma maior germinação em fluido de cirro diluído. Os componentes dos cirros reduziram a germinação a 10-60% de humidade relativa. Uma vez dispersos os esporos, os efeitos estimulantes do fluido dos cirros são provavelmente negligenciáveis (Griffiths e Peverett, 1980).
A 35-45% de humidade relativa, esporos de S. tritici em cirri permaneceu viável pelo menos 60 dias (Gough e Lee, 1985). Os componentes dos cirros podem actuar como inibidores da germinação dos esporos, ou o elevado potencial osmótico dos cirros pode impedir a germinação. Picnidiosporos de S. nodorum não sobreviveu durante 24 horas a uma humidade relativa superior a 80% a 20 C. Os esporos sobreviveram duas semanas ou mais a uma humidade relativa <10% (Griffiths e Peverett, 1980). Quando o fluido de cirrus de S. nodorum foi diluído com água, cerca de dois terços dos picnidiosporos perderam viabilidade em 8 horas, e após 30 horas à luz do dia, apenas 5% germinou. Quando os esporos foram armazenados no escuro, 40% permaneceu viável após 30 horas (Brennan et al., 1986).
Conídios secos de S. nodorum, à sombra e sob a luz solar directa, sobreviveu ao ar livre pelo menos 56 horas (Fernandes e Hendrix, 1986a). A germinação de S. nodorum pycnidiospores foi inibida por UV-B contínuo (280-320 nm), enquanto que a germinação de S. tritici não estava. A extensão do tubo de germes sob UV-B contínuo foi inibida para ambos os fungos, em comparação com a escuridão (Rasanayagam et al., 1995).
Infecção por Septoria nodorum
O processo de penetração do hospedeiro e desenvolvimento de S. nodorum dentro da folha foi examinada em pormenor por vários investigadores (Baker and Smith, 1978, Bird and Ride 1981, Karjalainen and Lounatmaa, 1986; Keon and Hargreaves, 1984; Straley, 1979; Weber, 1922). Os picnidiosporos tendem a alojar-se nas depressões entre duas células epidérmicas, e muitas tentativas de penetração das folhas começam aí. Os esporos germinam na superfície da folha em resposta à humidade livre (Fernandes e Hendrix, 1986b). Começam a germinar 2-3 horas após a deposição, e após 8 horas a germinação pode atingir 90%. A penetração das folhas começa cerca de 10 horas após a deposição dos esporos (Bird and Ride, 1981; Brönnimann et al., 1972; Holmes e Colhoun, 1974).
No início da germinação, o tubo germinal é rodeado por um material amorfo que se prende à folha. Os tubos germinativos que crescem de uma extremidade ou outra de um esporo e de células intercalares tendem a crescer ao longo das depressões entre células e são frequentemente orientados ao longo do longo eixo da folha (O'Reilly e Downes, 1986). As hifas de esporos não presentes nas depressões crescem aleatoriamente com ramificações ocasionais (Straley, 1979). Forma-se um appressorium com uma cavilha de infecção que penetra a cutícula e as paredes periclinais das células epidérmicas directamente no lúmen celular, resultando na morte rápida das células.
Muitas penetrações são primeiro o crescimento subcuticular ou lateral de uma hifa que ocorre dentro da parede celular antes do crescimento no citoplasma (Bird and Ride, 1981; O'Reilly and Downes, 1986). A penetração através de estomas abertos e fechados também ocorre e pode ser mais rápida que a penetração directa (Harrower, 1976; Jenkins, 1978; O'Reilly e Downes, 1986; Straley, 1979). Os tubos germinativos ramificam-se nos estomas e nas junções das células epidérmicas. A penetração de um tubo de germes num estômago pode ocorrer sem formação de um appressorium. A penetração ocorre por vezes através de tricomas (Straley, 1979). Aparentemente, a maioria das tentativas de penetração falham, com papilas densas formadas nas células no local da tentativa de penetração (Karjalainen e Lounatmaa, 1986; Bird and Ride, 1981).
Após a penetração, as células epidérmicas morrem rapidamente e tornam-se lenhificadas, e as hifas crescem para a mesofila. As células mesofílicas tornam-se deformadas, e o material lignificado é depositado fora de algumas células, que depois colapsam. A lenhificação ocorre antes de as hifas alcançarem a célula. O processo é o mesmo em todas as cultivares, mas desenvolve-se mais lentamente em cultivares resistentes. As hifas crescem intercelularmente entre as células epidérmicas, depois para a mesofila. Quando a mesofila é penetrada, a deterioração cloroplástica começa dentro de 6-9 dias (Karjalainen e Lounatmaa, 1986).
No entanto, a taxa fotossintética começa a diminuir dentro de um dia após a infecção e antes dos sintomas serem visíveis (Krupinsky et al, 1973). O tecido esclerênquima em redor dos feixes vasculares previne a infecção do tecido vascular. Os feixes vasculares bloqueiam a propagação de hifas através da mesofila, excepto quando o tecido de esclerênquima é jovem e não está totalmente formado (Baker e Smith, 1978).
O Stagonospora nodorum liberta uma vasta gama de enzimas degradantes da parede celular incluindo amilase, pectina metil esterase, poligalacturonases, xilanases, e celulase in vitro e durante a infecção de folhas de trigo (Baker, 1969; Lehtinen, 1993; Magro, 1984). A informação relacionada com a degradação da parede celular pelas enzimas concorda com as observações histológicas. Estas enzimas podem actuar em conjunto com toxinas. A sensibilidade enzimática pode estar relacionada com a resistência e taxa de colonização fúngica (Magro, 1984). Como muitos necrotrofos, os agentes patogénicos Septoria e Stagonospora produzem compostos fitotóxicos in vitro. A deterioração celular e a morte antes do crescimento hifálico em tecido mesofílico (Bird and Ride, 1981) é consistente com a produção de toxinas. Contudo, não foi estabelecido um papel definitivo para as toxinas no processo de infecção e a sua relação com a resistência do hospedeiro (Bethenod et al, 1982; Bousquet et al, 1980; Essad e Bousquet, 1981; King et al, 1983). As diferenças na gama de hospedeiros entre estirpes de trigo e cevada adaptadas à cevada são de S. nodorum pode estar relacionada com a produção de toxinas (Bousquet e Kollmann, 1998). O início da germinação dos esporos e a percentagem de esporos germinados não são influenciados pela susceptibilidade do hospedeiro (Bird and Ride, 1981; Morgan 1974; Straley, 1979; Straley and Scharen, 1979; Baker and Smith, 1978).
Bird and Ride (1981) relatou que a extensão dos tubos de germes na superfície da folha era mais lenta em cultivares resistentes do que em cultivares susceptíveis. Este mecanismo, expresso pelo menos 48 horas após a deposição de esporos, indica resistência de pré-penetração ao alongamento dos tubos germinativos. Houve menos penetrações bem sucedidas em cultivares resistentes, e a penetração prosseguiu mais lentamente em cultivares resistentes (Baker and Smith, 1978; Bird and Ride, 1981). A lenhificação foi proposta para limitar a infecção tanto em cultivares resistentes como susceptíveis, mas outros factores abrandaram o desenvolvimento fúngico em linhas resistentes. Nas linhas susceptíveis, a Hyphae de crescimento mais rápido pode escapar à lenhificação das células hospedeiras.Quatro dias após a inoculação da cevada com um biótipo de trigo isolado de S. nodorumAs hifas cresceram através da cutícula e por vezes nas camadas externas de celulose das paredes celulares epidérmicas. Papilas espessas foram depositadas sob as hifas de penetração e as células não foram penetradas (Keon e Hargreaves, 1984).
Infecção por Septoria passerinii: Green e Dickson (1957) apresentam uma descrição detalhada do processo de infecção de S. passerinii sobre a cevada. O processo de infecção é semelhante a S. tritici. Tal como S. tritici, o tempo necessário para a penetração das folhas é consideravelmente maior do que para S. nodorum. Os tubos germinativos ramificam-se e crescem sobre a superfície da folha ao acaso, mas por vezes ao longo de depressões entre as células epidérmicas. A penetração das folhas é quase exclusivamente através dos estomas. As hifas da germinação tornam-se inchadas, e se a penetração não for bem sucedida, as hifas continuam a alongar-se. Nenhuma penetração ocorre 48 horas após a deposição de esporos. Após 72 horas, os tubos de germes engrossam sobre os estomas, crescem entre as células de guarda e sobre as superfícies das células acessórias e para as cavidades sub-matais. A penetração directa entre as células epidérmicas só raramente é observada.
A germinação de esporos e a penetração do hospedeiro são as mesmas em cultivares resistentes e susceptíveis. Há muito menos extensão de hifas dentro das folhas em cultivares resistentes e as papilas são observadas em muitas mas não em todas as paredes celulares. As hifas crescem sob a epiderme de um estoma para outro, mas não penetram entre as células epidérmicas. A mesofila é colonizada, mas sem forma de haustoria. Após as células mesofílicas se tornarem necróticas, as células epidérmicas colapsam. O desenvolvimento micelial na folha é esparso e geralmente bloqueado por feixes vasculares. Nas folhas mais jovens, se a bainha vascular estiver menos desenvolvida, as hifas passam entre o feixe e a epiderme. Formam-se picnídeas em cavidades substomatais, principalmente na superfície superior da folha (Green e Dickson, 1957).
Factores que afectam a longevidade dos esporos sobre a superfície das folhas Entre os agentes patogénicos dos cereais Stagonospora e Septoria, só foram comunicadas informações definitivas sobre o processo de infecção para S. nodorum, S. triticie S. passerinii. Como muitos outros agentes patogénicos necrotróficos, nenhum dos grupos de agentes patogénicos suscita a reacção hipersensível. Uma diferença significativa no processo de infecção entre os agentes patogénicos Septoria e Stagonospora é que a germinação e a penetração dos esporos se processa muito mais rapidamente para S. nodorum do que para S. tritici e S. passerinii. Isto tem uma influência significativa na epidemiologia das doenças.
Os agentes patogénicos Septoria penetram na planta principalmente através dos estomas, enquanto S. nodorum penetra tanto directamente como através dos estômagos. S. nodorum penetra e mata as células epidérmicas rapidamente, mas S. tritici e S. passerinii não matar células epidérmicas até que as hifas tenham ramificado através da mesofila da folha e comece a necrose rápida. Os estudos histológicos do crescimento fúngico após a penetração do hospedeiro correspondem aos dados gerados a partir de estudos epidemiológicos de resistência do hospedeiro. A resistência abranda a taxa de colonização do hospedeiro, mas não tem um efeito apreciável no processo de desenvolvimento da lesão.
Os mecanismos de controlo da resposta do hospedeiro, quer relacionados com enzimas e toxinas ou outros metabolitos libertados pelos agentes patogénicos durante a infecção, ainda não estão claros. Há pouca informação sobre a infecção por ascósporos. O processo de infecção é provavelmente muito semelhante ao dos picnidiosporos. Os ascósporos de Phaeosphaeria nodorum germinam numa vasta gama de temperaturas, e os seus tubos germinativos penetram directamente na folha. No entanto, segundo Rapilly et al. (1973), os ascósporos, ao contrário dos picnidiosporos, não germinam em água livre.
Modelo de Infecção Septoria spp.
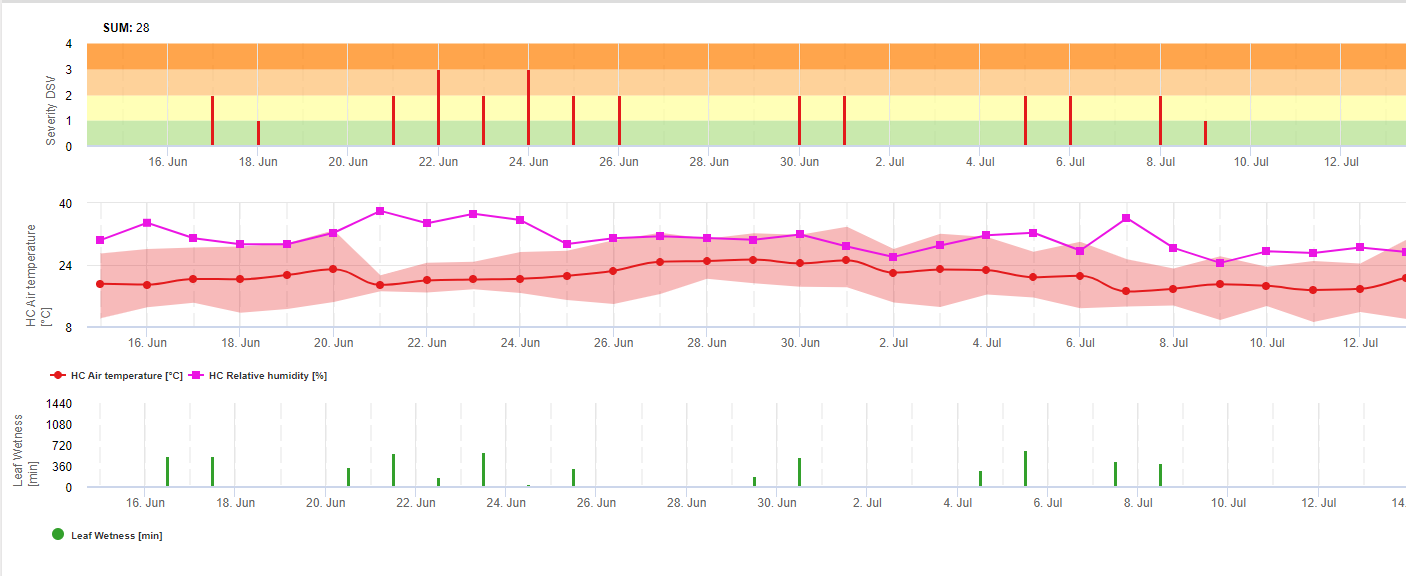
As infecções de Septoria são possíveis a baixas temperaturas enquanto que temperaturas inferiores a 7°C podem não levar a uma infecção no prazo de 2 dias. A temperatura óptima da doença é atingida na área de 16 a 21°C. As infecções são possíveis dentro de um período de elevada humidade relativa ou humidade foliar de 14 horas ou mais. Para satisfazer as condições, decidimos separar em modelos para infecções fracas, moderadas e graves. As infecções fracas podem ser dadas se for possível que o patogénico infecte o tecido hospedeiro. Isto significa que infecções fracas podem ter lugar se as temperaturas estiverem no mínimo e os períodos de humidade foliar forem de duração crítica. Uma infecção moderada terá lugar em condições em que a maioria dos ensaios de infecção conduza a resultados razoáveis e as infecções graves terão lugar em condições em que o agente patogénico tenha condições óptimas para a infecção.

Começa a infecção depois de uma chuva de 0,5 mm. Decidimos não utilizar um modelo para a formação de picnídeas. A condição necessária para a formação de picnídeas é suposta ser um período com humidade relativa superior a 85%. Espera-se que o tempo de vida das picnídeas seja de 24 horas. Em todos os climas onde a Septoria tritici tem a possibilidade de infectar, encontraremos 2 horas a cumprir esta condição quase todos os dias à volta do nascer do sol.
Avaliação da gravidade da infecção: Para podermos avaliar a pressão da infecção Septoria tritici entre a fase 10 (primeira folha através do coleóptero) e a fase 32 (nó dois pelo menos 2 cm acima do nó 1) e entre 32 e 51 (início do cabeçalho) temos de avaliar a gravidade das infecções com base nas condições climáticas. Esta avaliação é feita numa escala de 1 a 5. É dada uma severidade de 1 se a condição para uma infecção fraca estiver preenchida e tiver chovido menos de 5 mm, caso contrário, o vlaue de severidade correspondente será 2. É dada uma severidade de 3 se uma infecção moderada estiver preenchida e tiver chovido menos de 5 mm. Se tiver chovido mais de 5 mm durante uma infecção moderada ou menos de 5 mm durante uma infecção grave, é dada uma gravidade de 4.
Uma infecção grave com mais de 5 mm de chuva corresponde a um valor de gravidade de 5.
Septoria tritici avaliação da pressão da doença: O clima é apenas um dos factores que descem sobre a pressão da doença no campo. Os outros dois factores são a história do campo e a susceptibilidade da variedade cultivada. Se conseguirmos acumular os valores de gravidade da doença do estádio 10 ao estádio 32 até ao valor de 4, podemos esperar uma fraca pressão da doença pelo clima. Se este valor atingir 6, podemos esperar uma pressão moderada da doença e se atingir 10, podemos esperar uma pressão elevada da doença pelo clima. O conhecimento da susceptibilidade da variedade e da história do campo levar-nos-á a pulverizar ou não sobre uma pressão de doença fraca ou moderada nesta situação. Ter um valor acumulado de 10 pode levar a uma pulverização na fase 32 de qualquer forma. A decisão de uma pulverização numa fase posterior é mais dependente do clima da Primavera. Se formos capazes de acumular os valores de gravidade desde a fase 10 a um valor de 6, podemos esperar uma fraca pressão da doença. Se este valor atingir 10, podemos esperar uma pressão moderada da doença e se este valor atingir 15, podemos esperar uma pressão elevada da doença na situação climática.
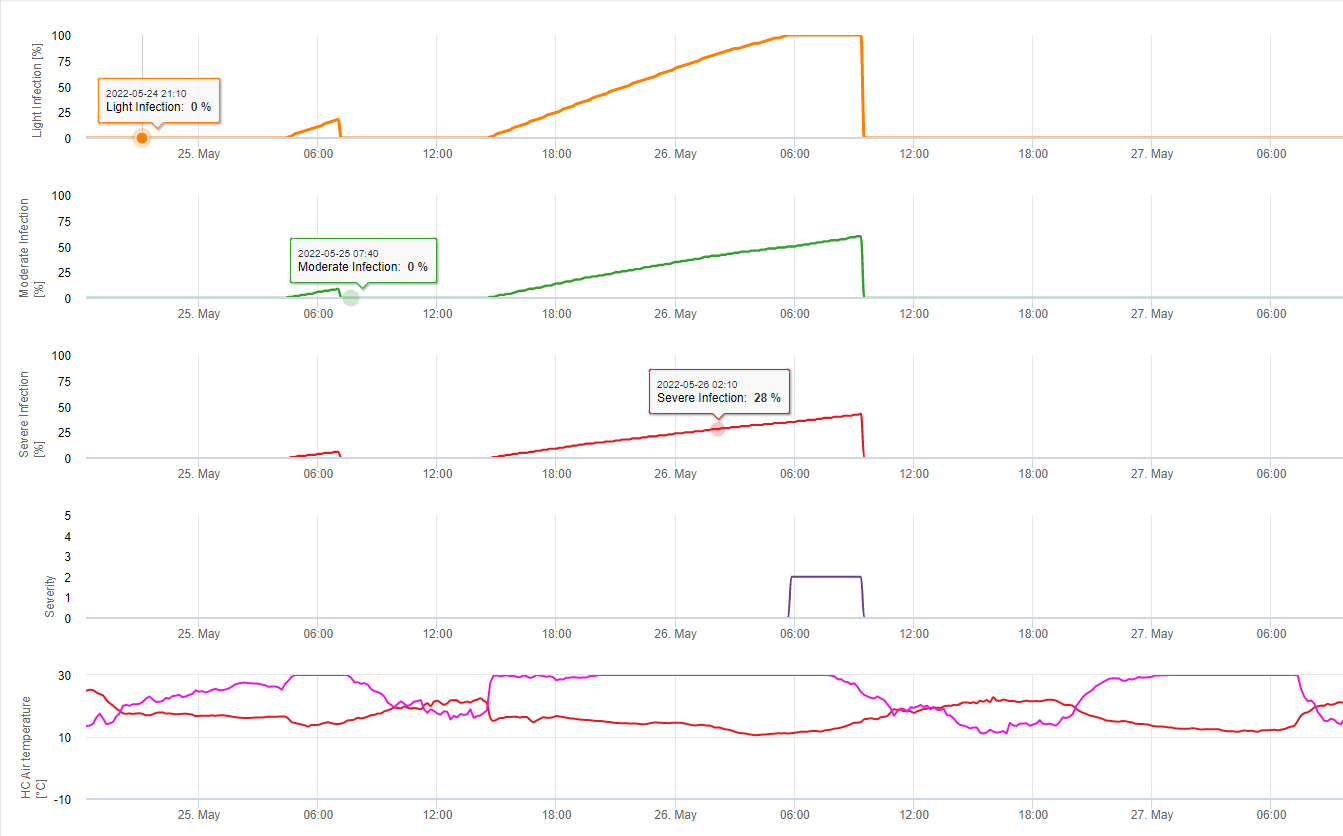
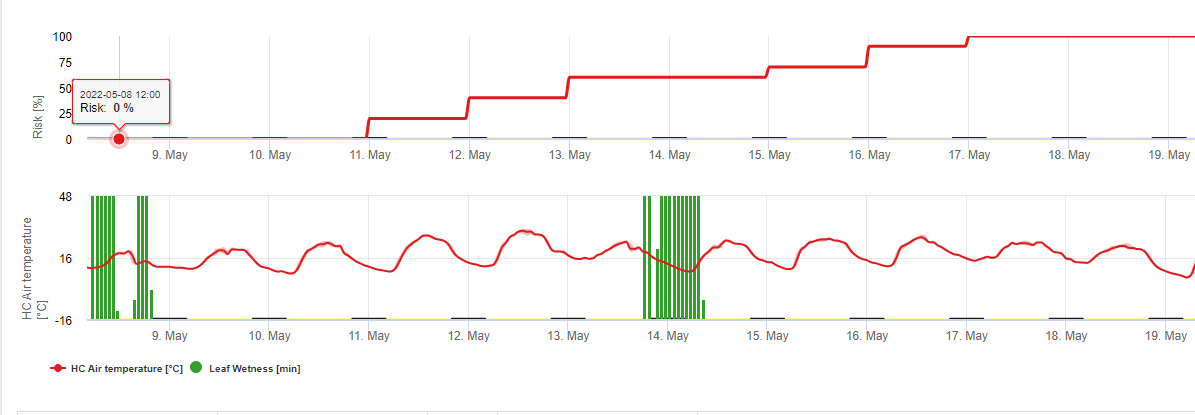
No FieldClimate mostramos o Septoria tritici Gravidade juntamente com as três gravidades diferentes de infecção num gráfico (ver acima). Devido a precipitação excessiva e longos períodos de humidade das folhas para uma infecção grave por S. tritici estiveram em pleno campo nos dias 14 e 16 de Maio. Os níveis de severidade atingem o valor mais alto de 5 em 14 de Maio, o que significa que um elevado risco de infecção é agora.
A biologia da infecção por Stagonospora nodorum difere de alguma forma de S. tritici, mas esta diferença não é suficientemente grande para um modelo separado. Por isso sugerimos a utilização deste modelo para todo o complexo de doenças de Stagnospora e Septoria em cereais, incluindo S. passerinii. S. tritici e S. passerinii tendem a precisar de períodos de molhamento das folhas mais longos do que S. nodorum. Em áreas com uma pressão elevada de S. nodorum As infecções classificadas a um fraco dando um valor de gravidade de 2 devem ser tratadas mais seriamente do que noutras áreas.
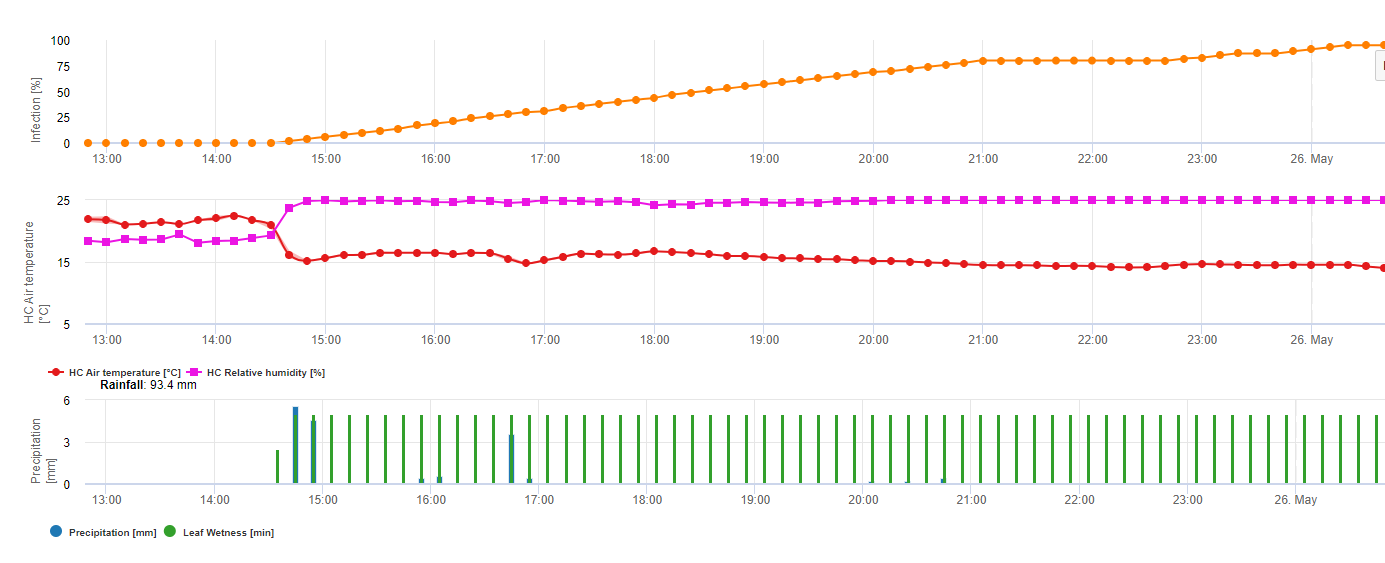
Para Septoria nodorum um modelo de risco é mostrado no FieldClimate (ver acima). Um risco elevado foi determinado nos dias 17 de Junho e 7 de Julho (100%). Dependendo da fase das plantas susceptíveis de infecção, as medidas de protecção das plantas têm de ser tidas em conta se o risco atingir 80% (ver também previsão do tempo do soro, protecção das plantas). Se o risco fosse 100% e já tivesse sido determinada uma infecção, as medidas sistémicas de protecção das plantas (aplicação curativa) têm de ser tomadas para proteger a planta.
Várias espécies de fungos patogénicos das plantas do género Colletotrichum causar antracnose em pimentos e muitos outros vegetais e frutas. Até finais dos anos 90, a antracnose em pimentos e tomates estava apenas associada a fruta madura ou amadurecida. Desde essa altura, estabeleceu-se uma forma mais agressiva da doença. Esta forma ataca o pimento em qualquer fase do desenvolvimento da fruta e pode ameaçar a rentabilidade das culturas do pimento nas áreas onde se estabelece. Esta doença pode também afectar o tomate, morangos, e possivelmente outras culturas fruteiras e hortícolas.
Sintomas
As lesões circulares ou angulares afundadas desenvolvem-se em frutos imaturos de qualquer tamanho. Muitas vezes formam-se lesões múltiplas em frutos individuais. Quando a doença é grave, as lesões podem coalescer. Muitas vezes, massas de esporos fúngicos rosa a laranja formam-se em anéis concêntricos na superfície das lesões. Em lesões mais antigas, podem observar-se estruturas negras denominadas acervuli. Com uma lente de mão, estas parecem pequenos pontos negros; sob um microscópio, parecem tufos de pequenos pêlos negros. O patogénio forma esporos rápida e profusamente e pode propagar-se rapidamente por uma cultura de pimenta, resultando em perda de rendimento até 100%. As lesões podem também aparecer em caules e folhas como manchas castanhas de forma irregular com bordos castanhos escuros.
Pathogen
Esta forma de antracnose de pimenta é causada pelo fungo Colletotrichum acutatum. O agente patogénico sobrevive em detritos vegetais de culturas infectadas e em outras espécies vegetais susceptíveis. O fungo não é transportado pelo solo durante longos períodos na ausência de detritos vegetais infestados. O fungo também pode ser introduzido numa cultura em sementes infestadas. Durante períodos quentes e húmidos, os esporos são salpicados pela chuva ou pela água da rega de fruta doente a fruta saudável. Os frutos doentes actuam como fonte de inóculo, permitindo que a doença se propague de planta para planta dentro do campo. O fungo sobrevive dentro e sobre as sementes. A antracnose é introduzida no campo em transplantes infectados ou pode sobreviver entre estações em detritos vegetais ou em hospedeiros de ervas daninhas. Os hospedeiros alternativos incluem ervas daninhas e outras plantas em Solanaceae (tomateiro, batata, beringela) embora as infecções destes hospedeiros sejam extremamente raras na Florida. Os frutos são infectados quando esporos do fungo ou detritos infestados são salpicados pela chuva sobre plantas pimenteiras. Novos esporos são produzidos dentro do tecido infectado e depois são dispersos por outros frutos. Os trabalhadores podem também deslocar esporos com equipamento ou durante a manipulação de plantas infectadas. A infecção ocorre geralmente durante o tempo quente e húmido. Temperaturas à volta de 80° F (27° C) são temperaturas óptimas para o desenvolvimento de doenças, embora a infecção ocorra tanto a temperaturas mais altas como a temperaturas mais baixas. Perdas graves ocorrem durante o tempo chuvoso porque os esporos são lavados ou salpicados para outros frutos, resultando em mais infecções. É mais provável que a doença se desenvolva em frutos maduros que estejam presentes durante um longo período na planta, embora possa ocorrer tanto em frutos imaturos como em frutos maduros. A antracnose pode infectar de 15 °C a 30 °C. Mas uma humidade das folhas longas é necessário para satisfazer as necessidades de uma infecção. À temperatura óptima de 20°C a 25°C ainda 12 horas de humidade das folhas são necessárias. Uma temperatura mais elevada ou mais fria necessitará de períodos de chicotada das folhas ainda mais longos (sem função linear/arranjo necessário para o cálculo). FieldClimate calcula os possíveis eventos de infecção com base na humidade das folhas e as temperaturas durante este evento.
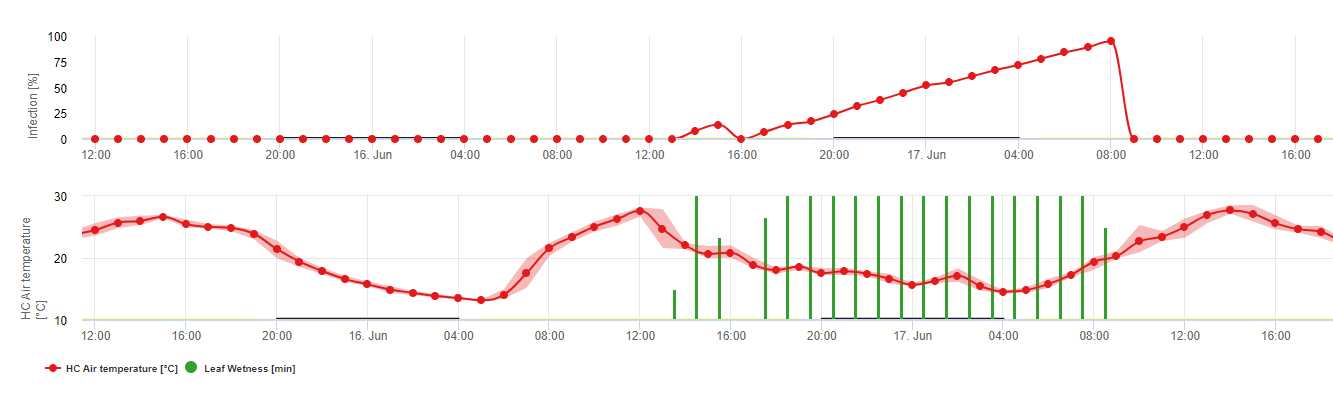
Biologia de Plasmopara viticola (Downy Mildew)
P. viticola é um parasita obrigatório. O que significa que órgãos verdes e frescos da vinha são necessários para crescer. Durante o período sem vegetação persiste a formação de corpos frutíferos, os chamados oósporos. Os oósporos de oomycetes podem sobreviver a períodos muito longos no solo. Portanto, podemos encontrar míldio em locais onde as infecções não são possíveis em cada ano. Na primavera, quando o solo superior estiver suficientemente húmido e quente, os oósporos formarão a chamada macrosporangia, que pode libertar até 200 zoósporos em água livre. Os zoósporos movem-se até às folhas e aglomerados pelo vento em gotículas de água. Têm dois flagelos e movem-se numa película de água no lado negativo das folhas ou dos cachos e bagas jovens para encontrar um estoma para entrar no tecido vegetal. Entram e germinam no estoma, no qual transferem todo o seu plasma em menos de uma hora. Em estudos microscópicos, a descoberta do estoma, a encistação e a germinação para o estoma foi concluída em 90 minutos.
P. viticola cresce no espaço intercelular e alimenta-se de hemostorias que penetram as células epidérmicas e parenquimatosas. Em dependência da temperatura e humidade relativa, desenvolve um crescimento intercelular suficiente com suficientes haustorias para formar um corpo de substrato que preenche toda a área do substrato e que levanta o tecido epidérmico do tecido parenquimatoso. Isto leva ao sintoma visível da mancha de óleo.
Oomycetes são esporulando na ausência de luz quando a humidade relativa é muito elevado. Em P. viticola não há sporulação se as temperaturas forem inferiores a 12°C e a humidade relativa for inferior a 95%. Os esporangiaforos são formados pelas vesículas do substrato e sairão do estoma. Os esporângios recém-formados são pegajosos e só podem ser removidos dos esporângios pela água. Durante a diminuição da humidade relativa, os esporângios tornam-se tentativa e também podem ser removidos pelo vento.
Os esporângios libertarão até 20 zoosporos em água livre. Estes zoosporos também têm de ser distribuídos pelo vento em gotículas de água para chegarem às folhas frescas, ou os esporângios podem ser distribuídos pela chuva ou pelo próprio vento. O processo de infecção da infecção primária e secundária é o mesmo.
Fazer à grande importância da fase sexual para a hibernação do agente patogénico, podemos assumir tipos de acasalamento adequados em todas as vinhas onde ocorre míldio da videira. A formação de zoósporos tem lugar em folhas mais velhas durante o Verão e no início do Outono.
Infecção primária em detalhe
As infecções provenientes dos oósporos são chamadas infecções primárias. Este termo é enganador, uma vez que várias infecções primárias podem ocorrer no início do Verão. Em eventos epidemiológicos, a infecção primária não desempenha um papel importante se houver manchas de óleo suficientes nas vinhas e o potencial de infecção dos esporos de Verão (esporângios) exceder os esporos.
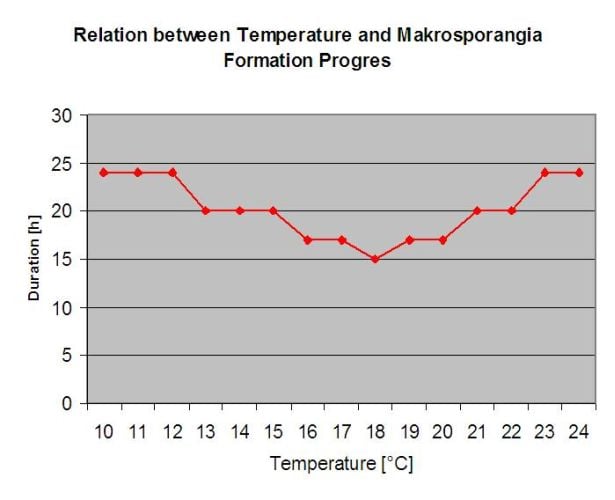
Os oósporos de Inverno desenvolvem-se para os chamados macrosporangia sempre que há humidade relativa suficiente para cerca de 24 horas. Os macrosporângios libertam os seus zoosporos em água livre e chuva intensa (como numa trovoada). Os esporos são transportados para as folhas das videiras e/ou rebentos verdes. Infecções primárias, portanto, precisa de mais tempo de chuva períodos ou várias trovoadas sucessivas. A primeira chuva faz com que as folhas caídas sejam saturadas com água e a chuva forte nos dias seguintes faz com que as macrosporângios libertem os seus zoospórdios, que atingem as folhas ou rebentos da videira. Para o sucesso da infecção, é necessário um período suficientemente longo de humidade das folhas para permitir que os zoósporos alcancem o estoma das folhas ou rebentos e os infectem (a Ilustração acima mostra o ciclo de desenvolvimento do míldio da videira (Plasmopara viticola).
O Modelo para o Downy Mildew verifica primeiro, se o tempo é adequado para o desenvolvimento de macrosporangia. Este é o caso desde que as folhas estejam molhadas, ou que a humidade relativa após a chuva não desça abaixo de 70%. Dependendo da temperatura, as macrosporângios podem desenvolver-se dentro de 16 a 24 horas. Se as macrosporângias maduras estiverem disponíveis, serão mostradas graficamente na exibição da infecção primária por Downy Mildew. Se as macrosporângios estiverem presentes, uma forte precipitação pode espalhar os seus zoospóros. Uma chuva contínua de 5 mm é interpretada como uma chuva forte e os zoósporos são espalhados, uma infecção primária é mais do que possível se a humidade das folhas tiver durado o tempo suficiente.
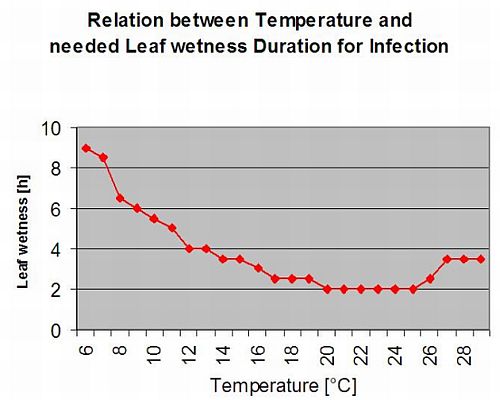
Infecções Secundárias
Infecções secundárias de Plasmopara viticola só são possíveis se já existirem manchas de óleo maduras na sua vinha ou perto dela. As manchas de óleo estão maduras quando são capazes de esporângio (produzir esporângios). Os esporângios só são produzidos por noite. A luz solar inibe a esporulação. Os esporângios são produzidos se for mais quente que 12°C e a humidade relativa é muito elevado. A taxa de produção de esporângios aumenta com a temperatura até 24°C. A temperatura óptima para o esporângio nas castas europeias (Vitis vinifera) é de cerca de 24°C. Se as temperaturas excederem os 29°C, não se pode fazer qualquer esporulação. No nosso modelo, verificamos se humidade de mais de 95% ocorre durante a noite. Se esta condição durar uma temperatura horária acumulada de mais de 50°C, a esporulação terminou e novos esporângios de Plasmopara existem na vinha. Por exemplo, 50°C horas correspondem a 4 horas com 13°C ou 3 horas com 17°C.
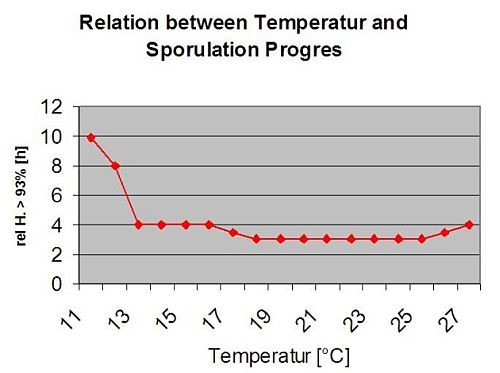
O gráfico seguinte mostra a relação entre as condições de temperatura e humidade (humidade relativa, humidade foliar, precipitação) que são utilizadas para modelar os eventos de infecção em fieldclimate.com.
O oídio do tomate pode ser causado por três agentes patogénicos em todo o mundo.
Leveillula taurica (Oidiopsis taurica) é um agente patogénico de uma vasta gama de espécies hospedeiras em climas áridos a semi-áridos quentes na Ásia, Mediterrâneo, África, e mais recentemente no sudoeste dos Estados Unidos.
Erysiphe orontii (E. cichoracearum e E. Polifaga) é outra espécie comum a muitas plantas hospedeiras, tanto em regiões temperadas como tropicais.
E como uma terceira espécie Odium lycopersicum.
Factores para o Desenvolvimento de Doenças
- níveis de humidade relativa > 50% (humidade relativa óptima > 90%)
- não é necessária água livre nas superfícies foliares
- gama de temperaturas: 10-35 °C (melhor abaixo de 30 °C)
O míldio em pó é uma doença induzida por inóculos. Por conseguinte, apenas podem ser determinados períodos de risco, factor principal para os danos, por exemplo, o surto é o inóculo inicial (que está activo numa vasta gama de temperaturas). Assim, para as cintas de controlo: combinar a modelação do período de risco com a monitorização do inóculo fúngico (doença) no campo!
Tomate de campo aberto
Ferrugem dos espargos Puccinia asparagi é favorecido por um clima húmido e quente. Pequenas quantidades de chuva e vento distribuem os urediaspores, enquanto as chuvas fortes podem lavá-los até ao solo. Por esta razão, um período de humidade das folhas com mais de 10 mm de chuva não é tomado para um período de infecção. Os períodos de chuvas leves e orvalho de 3 a 8 horas com temperaturas de 25°C até 10°C podem ser períodos de infecção para os urediasporos no Verão e no final do Verão.
Este modelo estima a data da primeira ocorrência de C. beticola com base nas temperaturas médias diárias acumuladas desde o dia 1 de Janeiro acima dos 5°C. É necessário um limiar de 1006,2°C em períodos com humidade relativa superior a 60% e se não tivermos humidade relativa, é válido um limiar de 1081,9°C.
Este modelo foi desenvolvido para a Itália e é utilizado em Itália e na Alemanha.
Para o ano 2010 em Styria CercoPrim indica o 19 de Junho para o primeiro spray. Esta é aproximadamente a mesma data que o modelo de risco Pessl Instruments ou o modelo DIV teria indicado para aspergir variedades susceptíveis.
O Pathogen
A doença da mancha roxa em lanças de espargos e samambaia é causada pelo fungo Estemphylium vesicarium. O fungo sobrevive ao inverno como esporos sexuais (ascósporos) num saco (ascus) produzido em estruturas de Inverno (pseudotecia) que aparecem como pequenos pontos negros em detritos de espargos da estação anterior. O os ascósporos são libertados do ascus pela chuva e podem ser transportados pelo vento para as plantas de espargos recentemente surgidas, onde causam a infecção primária da estação de crescimento. Estas novas infecções resultam em esporos (conidia) produzidos por um processo assexuado, que por sua vez podem causar infecções secundárias, um processo que se repete desde que as temperaturas e a pluviosidade sejam favoráveis.
A Doença
O aparecimento da mancha roxa como um problema significativo na produção de espargos no Michigan pode dever-se à adopção de um sistema cultural "no till till", em que o feto seco da estação anterior é cortado em Abril e deixado na superfície do solo. Este resíduo persiste durante a época de colheita (meados de Junho) e é visível durante o período de crescimento da samambaia (finais de Junho a Setembro) e é a fonte de ascósporos que iniciam infecções no início da época de crescimento. Os sintomas da doença são: A doença aparece como numerosas manchas arroxeadas, ligeiramente afundadas, com centros castanhos que ocorrem em lanças e samambaias colhidas. As lesões nas lanças são frequentemente encontradas no lado do barlavento, porque o sopro de areia provoca feridas que favorecem a infecção. Durante anos epidémicos, a mancha pode ocorrer em 60-90% das lanças e pode resultar na rejeição da cultura, especialmente para a venda de produtos frescos no mercado. Também ocorrem manchas nos fetos dos espargos, afectando o caule principal, ramos secundários e agulhas (cladophylls). Uma infecção grave da samambaia pode resultar na desfoliação prematura da planta. O aumento da gravidade da doença da mancha roxa está associado a longos períodos de chuvas, nevoeiro ou orvalho.
Crédito de texto: Mary K. Hausbeck, Professora e Especialista em Extensão, Michigan State University, Department of Plant Pathology, E. Lansing
TomCast para Espargos
Antecedentes: O TOMCAST (TOMato disease foreCASTing) é um modelo informático baseado em dados de campo que tenta prever o desenvolvimento de doenças fúngicas, nomeadamente Early Blight, Septoria Leaf Spot e Anthracnose em tomates. Os registadores de dados colocados no campo estão a registar dados horários de humidade das folhas e de temperatura. Estes dados são analisados durante um período de 24 horas e podem resultar na formação de um Valor de Gravidade da Doença (DSV); essencialmente um incremento do desenvolvimento da doença. À medida que o DSV se acumula, a pressão da doença continua a aumentar sobre a cultura. Quando o número de DSV acumulado excede o intervalo de pulverização, recomenda-se uma aplicação de fungicida para aliviar a pressão da doença.
TOMCAST é derivado do modelo original F.A.S.T. (Forecasting Alternaria solani on Tomatoes) desenvolvido pelo Dr. Madden, Pennypacker, e MacNab na Universidade Estadual da Pensilvânia (PSU). O modelo PSU F.A.S.T. foi ainda modificado pelo Dr. Pitblado no Ridgetown College em Ontário, no que agora reconhecemos como o modelo TOMCAST utilizado pela Ohio State University Extension.
DSV são: Um Valor de Gravidade da Doença (DSV) é a unidade de medida dada a um incremento específico do desenvolvimento da doença (Early Blight).
Por outras palavras, um DSV é uma representação numérica de quão rápida ou lenta a doença (Early Blight) se está a acumular num campo de tomate. O DSV é determinado por dois factores: humidade da folha e temperatura durante as horas de "molho da folha". À medida que o número de horas de folha molhada e a temperatura aumenta, o DSV acumula a um ritmo mais rápido. Ver a Tabela de Valores de Gravidade da Doença abaixo.
Inversamente, quando há menos horas húmidas de folhas e a temperatura é mais baixa, o DSV acumula-se lentamente, se é que se acumula. Quando o número total de DSV acumulados excede um limite predefinido, chamado intervalo ou limiar de pulverização, recomenda-se um spray fungicida para proteger a folhagem e a fruta do desenvolvimento de doenças.
O intervalo de pulverização (que determina quando se deve pulverizar) pode variar entre 15-20 DSV. O DSV exacto que um produtor deve utilizar é normalmente fornecido pelo processador e depende da qualidade da fruta e da utilização final dos tomates. Seguindo um intervalo de 15 DSV spray é uma utilização conservadora do sistema TOMCAST, o que significa que pulverizará mais frequentemente do que um cultivador que utiliza um intervalo de 19 DSV spray com o sistema TOMCAST. A troca está no número de pulverizações aplicadas durante a estação e no potencial de diferença na qualidade da fruta.
 TomCast com Espargos
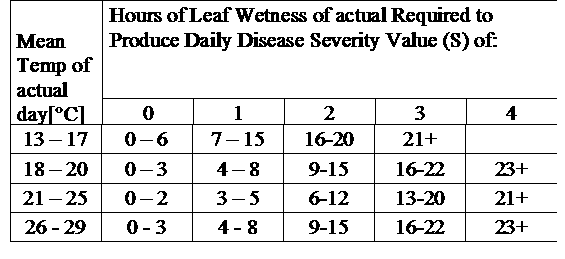
TomCast com Espargos
A investigação determinou que o Tom-Cast Disease forecaster é uma alternativa promissora à pulverização de samambaia em campos comerciais de espargos com base em calendário. O Tom-Cast alerta os cultivadores quando o as condições ambientais são favoráveis ao desenvolvimento da doença da mancha roxa (períodos prolongados de orvalho ou de chuva acompanhados de temperaturas quentes). Os fungicidas eficazes aplicados de acordo com a previsão da doença de Tom-Cast permitem aos cultivadores gerir a doença da mancha roxa dos espargos, poupando dinheiro e preservando o ambiente.
Crédito de texto: Jim Jasinski, Coordenador de TOMCAST PARA OHIO, INDIANA, & MICHIGAN
Modelo de infecção por manchas roxas
O Stemphylium versicarium modelo de infecção baseia-se no trabalho de SUHERI e PRICE em cebolas e LLorent, VILARDELLL, BUGIANI, GHERARDI e MONTESINO em pêra.
As curvas de infecção para infecções leves, moderadas e graves são computadas.
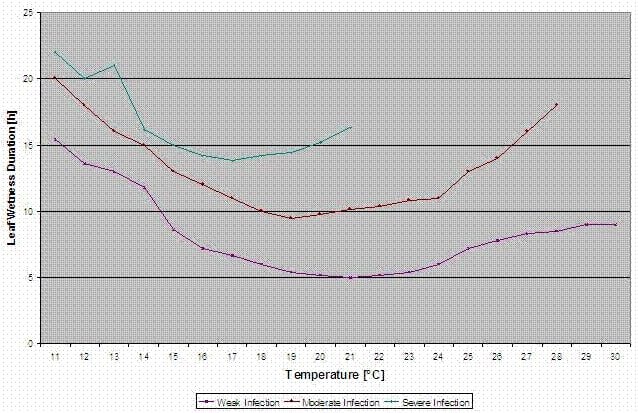
Com esta separação em situações favoráveis, moderadamente favoráveis e muito favoráveis, cabe ao cultivador decidir quão grande será a pressão da doença num campo específico e se ele tem de cobrir uma infecção específica.
Condições:
Temperatura: 11-30°C
Humidade da folha > 0 (início para infecção) ou humidade rel. > 90% (apenas se já havia uma infecção antes calculada e ainda está a aguentar)
infecção leve (11-30°C), infecção moderada (13-30°C), infecção severa (16-30°C), uma matriz com duração de molhamento das folhas (ver acima)
Repor: se a humidade rel. for inferior a 70%.