

Kórokozó
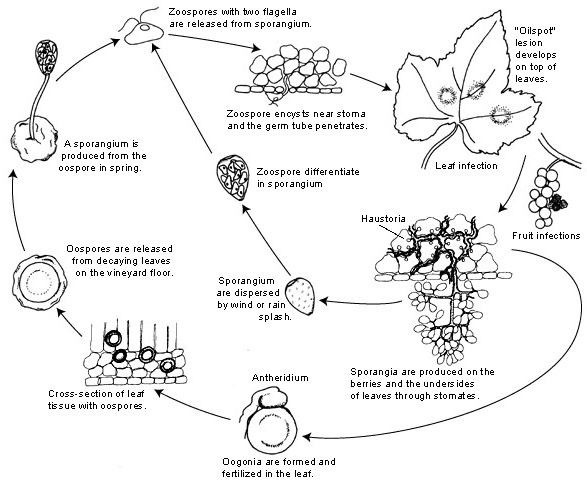
A szőlő lisztharmat kórokozója a következő Plasmopara viticola. Kötelező parazita, ami azt jelenti, hogy a növekedéshez zöld és friss szőlőszervekre van szükség. Aszexuális és ivaros szaporodási stádiuma is van - zoospórák és oospórák. Ezért a fertőzésnek két szakasza van: elsődleges és másodlagos fertőzés.
Az elsődleges fertőzés a telelő oospórák csírázásával kezdődik, amelyek tavasszal, amikor az időjárás felmelegszik, sporangiumokat képeznek. A sporangiumok éjszaka termelődnek, mivel a napfény gátolja a sporulációt. A vastag falú oospórák segítenek a kórokozónak túlélni a zord téli körülményeket és elindítani az elsődleges fertőzést. A sporangiumokat eső vagy szél szórja szét, és a szabad vízben zoospórákat bocsátanak ki. A sikeres fertőzéshez kellően hosszú levélnedvesség vagy esőzés szükséges.
A másodlagos fertőzés csak érett olajfoltok jelenlétében lehetséges, és zoosporák és sporangiumok útján történik. A zoospórák és sporangiumok rendkívül érzékenyek az alacsony páratartalomra és a fényre, ami lerövidíti életképességüket, ami azt jelenti, hogy a fertőzés nagy része hamarosan a felszabadulás után történik. A zoospórák csíracsöveken keresztül behatolnak a növényi szövetekbe, és a további fertőzéshez hifákat fejlesztenek. Ezek a hifák a hőmérséklettől függően 5 és 21 nap közötti lappangási idő után olajos elváltozásokat képeznek - minél alacsonyabb a hőmérséklet, annál tovább tart.
Ahogy az időjárás ismét hidegebbé válik, ami kedvezőtlen a kórokozó számára, a kórokozó áttér az ivaros termelésre, és oospórákat termel. Ezek az oospórák télen nyugalmi állapotban maradnak, és a ciklus ismétlődik.
Tünetek

A levelek felső felületén apró sárga sérülések jelennek meg, amelyeket néha barnás fényes foltok vesznek körül. Az olajfoltok az érés során kiterjednek és integrálódnak, a halók pedig elhalványulnak. A sporangiumok később a sérülések alján fehéres-tövises lisztharmatot hoznak létre, majd elhalás következik. A hajtásokon, szárakon és bogyókon is megjelennek olajos barna területek. Meleg, párás éjszakákon a spóraszaporodás következtében fehér pelyvával boríthatók. A súlyos fertőzés idő előtti lehullást okoz.
FieldClimate modellek
- Az FieldClimate-ben két modell létezik - elsődleges és másodlagos fertőzés.
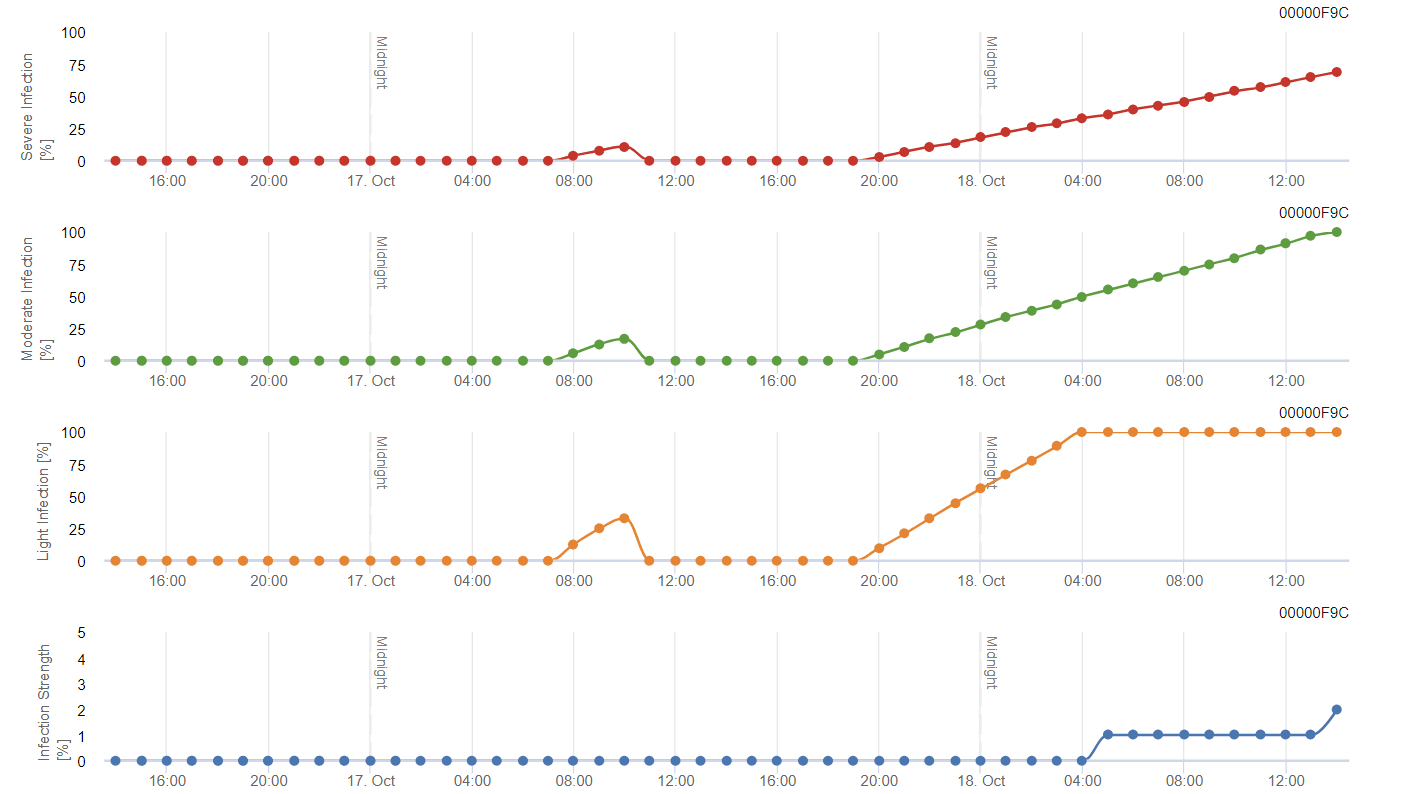
- Az első grafikon a fertőzés befejezése utáni lappangási időt mutatja. Az alsó grafikonok a fertőzés előrehaladását mutatják gyenge, közepes és súlyos fokozat esetén. Fertőzésről akkor kell feltételezni, ha a grafikonon az emelkedő görbe eléri az 100% értéket - a permetezés akkor jöhet szóba, ha a fertőzés súlyos szintje mutatkozik.
Elsődleges fertőzés
Szükséges érzékelők:
- Levegő hőmérséklete
- Relatív páratartalom
- Levélnedvesség
- Csapadék
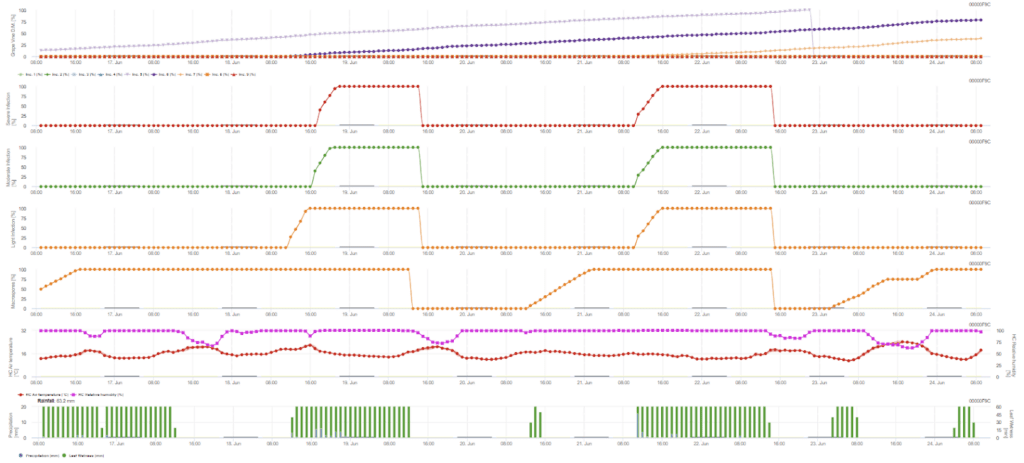
Az elsődleges fertőzés azt ellenőrzi, hogy az időjárás alkalmas-e a sporangiumok kialakulására. Ez mindaddig így van, amíg a levelek nedvesek, vagy az eső utáni relatív páratartalom nem csökken 70% alá. A sporangiumok a hőmérséklettől függően 16-24 órán belül fejlődhetnek ki. Az 5 mm-es folyamatos eső olyan erős esőzésként értelmezhető, amely a zoospórákat terjesztheti.
Másodlagos fertőzés
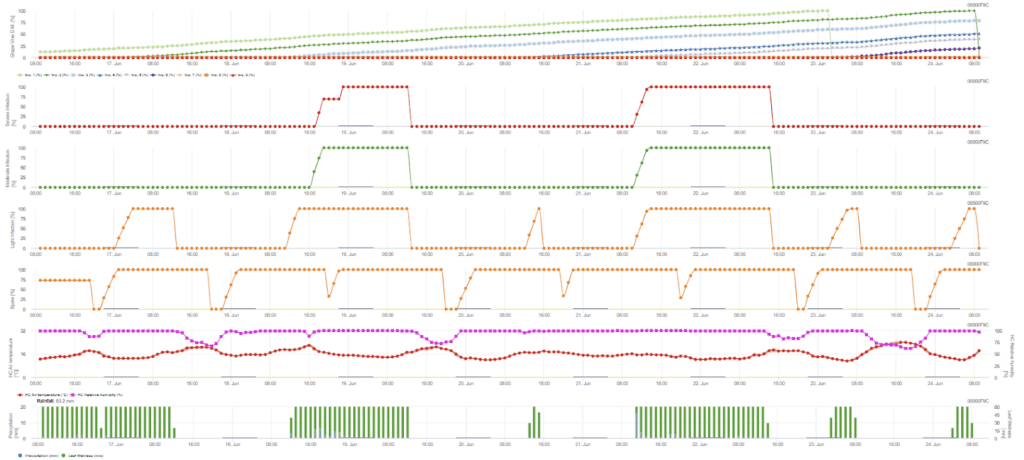
Szükséges érzékelők:
- Levegő hőmérséklete
- Relatív páratartalom
- Levélnedvesség
- Csapadék
A másodlagos fertőzés ellenőrzése akkor történik, ha a hőmérséklet 12 °C-nál melegebb és a relatív páratartalom 95% felett van. A sporangiumtermelés mértéke 23 °C-ig emelkedik a hőmérsékleten. Ha ez a feltétel 50 °C-nál magasabb óránkénti hőmérsékleten tart, akkor feltételezzük, hogy a sporuláció befejeződött, és új sporangiumok vannak a szőlőben. A felhalmozott 50 °C például 13 °C esetén 4 órának, 17 °C esetén 3 órának felel meg. A sporangiumok gyorsan elpusztulnak, ha melegebb és szárazabb lesz - ha a relatív páratartalom 50% alá csökken, 0-ra állítjuk vissza, és ha a hőmérséklet meghaladja a 29 °C-ot, akkor már nem történhet sporuláció.
Irodalom
- Ash, G. (2000). A szőlő lisztharmata. The Plant Health Instructor.
- Gessler, C., Pertot, I., & Perazzolli, M. (2011). Plasmopara viticola: a szőlő lisztharmatával kapcsolatos ismeretek áttekintése és a betegség hatékony kezelése. Phytopathologia Mediterranea, 50(1), 3-44.
- Kennelly, M. M., Gadoury, D. M., Wilcox, W. F., Magarey, P. A., & Seem, R. C. (2007). A szőlő lisztharmat kórokozója, a Plasmopara viticola primer fertőzése, a léziók produktivitása és a sporangiumok túlélése. Phytopathology, 97(4), 512-522.
- Koledenkova, K., Esmaeel, Q., Jacquard, C., Nowak, J., Clément, C., & Ait Barka, E. (2022). Plasmopara viticola a szőlő lisztharmat kórokozója: a taxonómiától a betegség kezeléséig. Frontiers in Microbiology, 13, 889472.
Kórokozó
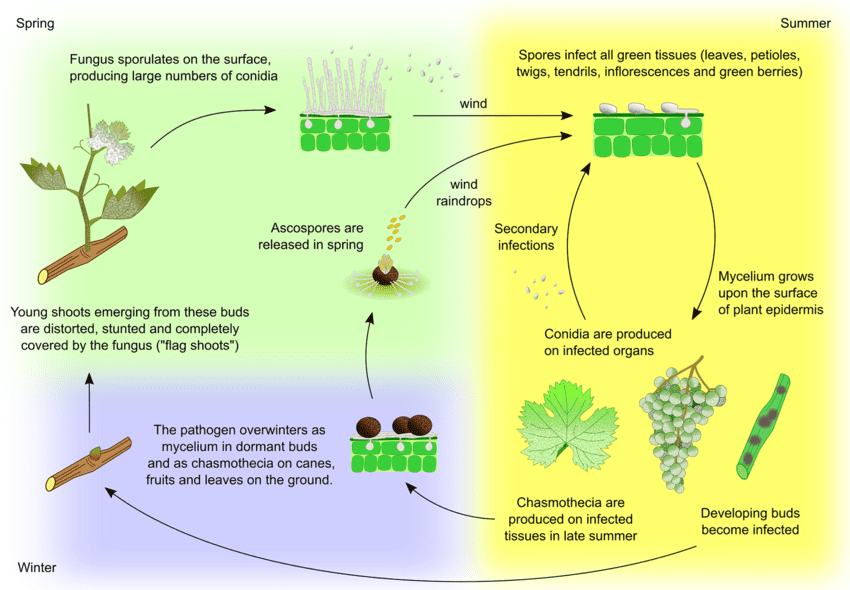
A szőlő lisztharmatának kórokozója az Uncinula necator. Az inokulumnak két fő forrása van - a telelő micélium és a kleisztotéciumokból származó aszkospórák -, amelyek jelentősége a régiótól függően változhat.
A micélium tavasszal zászlós hajtásokat okoz. A zászlós hajtások részben vagy teljesen lisztharmattal borított hajtások, amelyek látens fertőzött rügyekből képződnek. Mivel belül kolóniák képződnek, ezek a hajtások jól láthatók, és gyakran torz levelek is vannak rajtuk.
A kleisztotéciumok kora tavasszal aszkospórákat bocsátanak ki. A csapadék fontos tényező az aszkospórák kibocsátásában. A hőmérséklet emelkedésével a dehiszkozitás gyakrabban fordul elő. Az aszkospórák 12 órán belül kicsíráznak, csíracsöveket és appresszóriumot képeznek a növényi szöveteken, ami szétszórt kolóniaképződéshez vezet.
A másodlagos fertőzés a telepekben termelődő konídiumok útján történik. Az aszkospórákhoz hasonlóan ezek is kicsíráznak, csírázócsöveket és appresszóriumot fejlesztenek. Az U. necatornak nincs szüksége szabad vízre a fertőzéshez és magas relatív páratartalomra a konídiumképződéshez. Ha az időjárási körülmények kedvezőtlenebbé válnak, a gomba kleisztotéciumokat képez. A gombafaj a kleisztotéciumokban vagy vegetatív micéliumként telel át a nyugalmi állapotban lévő fertőzött rügyekben, és a ciklus ismétlődik.
Tünetek

A tünetek a szőlő minden zöld részén megtalálhatók. A hajtásokon telepek fejlődnek, és fehér-szürkés micéliummal borított, úgynevezett "zászlós hajtások" jelennek meg. Torz leveleket hoznak létre. A leveleken is találhatók lisztharmat telepek - akár az alsó, akár mindkét oldalon. A bogyókon hamuszürke foltok keletkeznek, és súlyos esetben felszakadnak. A vesszők sötétvörös-barna foltokat mutatnak, és a csúcsoktól kezdve elhalhatnak.
FieldClimate modellek
Ascospórák fertőzése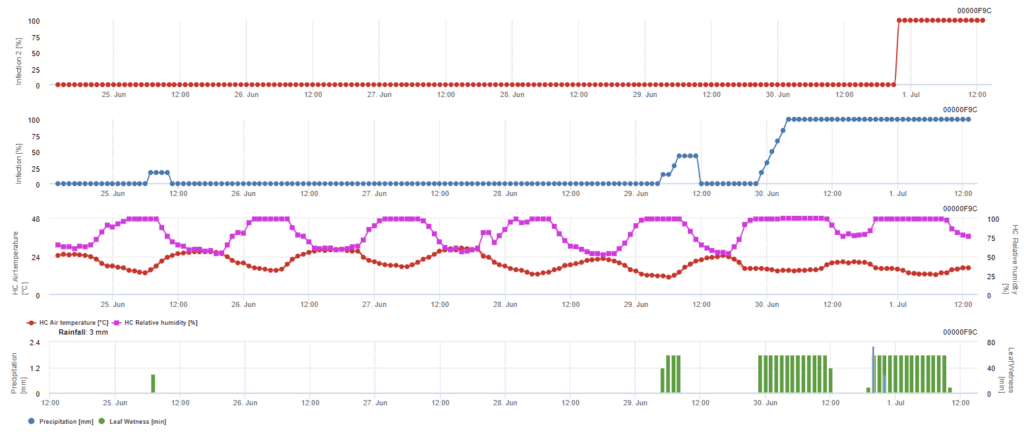
Szükséges érzékelők:
- Levegő hőmérséklete
- Relatív páratartalom
- Levél nedvesség
- Csapadék
Ez a modell akkor ajánlott, ha az aszkospórák a hideg téli időszakban képződnek (tehát nem a micéliummal fertőznek). Az aszkospórás fertőzés az aszkospórák felszabadulását és a kezdeti fertőzést a hosszabb levélnedves időszakok alatti átlaghőmérsékletek alapján jósolja meg. A számításhoz körülbelül 2,5 mm csapadék szükséges az aszkospórák felszabadulásához, amelyet legalább 8-12 órás levélnedvesség és 10-15 °C közötti hőmérséklet követ. A fertőzés bekövetkezése után a modell átvált a betegség kockázatának felmérési fázisára (kaliforniai kockázati modell), és a hőmérsékletnek a kórokozó szaporodási sebességére gyakorolt hatásán alapul.
Kaliforniai kockázati modell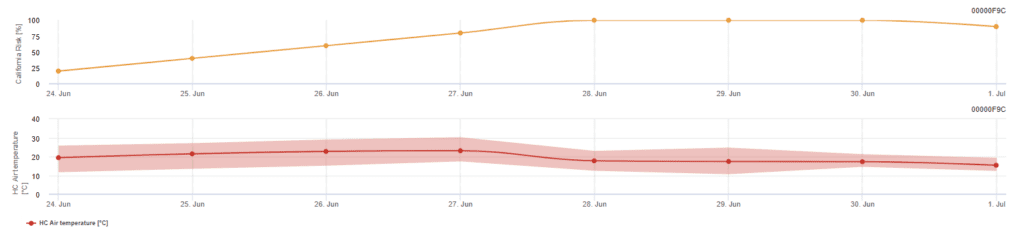
Szükséges érzékelők:
- Levegő hőmérséklete
A modell kaliforniai laboratóriumi vizsgálatokon alapul. Az aszkospórák felszabadulását és csírázását (modell) követően a lisztharmat későbbi fejlődését és szaporodását a hőmérséklet befolyásolja. A modell értékeli a hőmérsékleteket, és egy 0 és 100 pont közötti index segítségével értékeli a lisztharmat kialakulásának kockázatát.
A kockázatértékelési index elindításához három egymást követő napon legalább hat egymást követő órán át 21 és 30'c közötti hőmérséklet szükséges. Minden olyan napért, amikor 6 vagy több egymást követő órában 21 és 32 °C közötti hőmérsékletet teljesít, 20 pontot kap, és 10 pontot veszít azokért, amikor ez nem teljesül, vagy amikor a hőmérséklet meghaladja a 32 °C-ot, vagy 21 °C alá csökken.
Az alacsony, 0~30-as indexértékek azt jelzik, hogy a kórokozó nem szaporodik. A 40~50-es index közepesnek tekinthető, és körülbelül 15 napos szaporodási időt jelent a lisztharmat esetében. A 60 feletti indexértékek azt jelzik, hogy a kórokozó gyorsan (5 naponta) szaporodik, és nagy a járványos betegség kialakulásának kockázata.
Pessl Instruments kockázati modell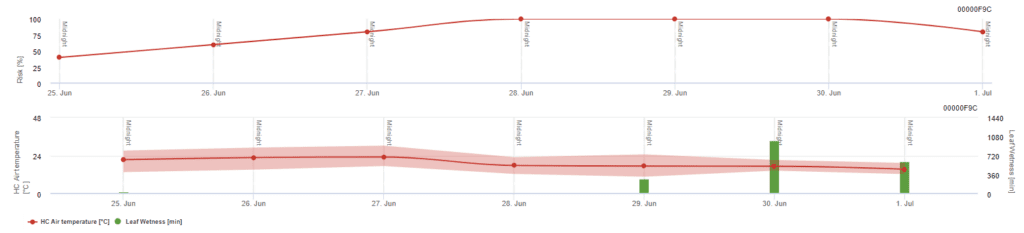
Szükséges érzékelők:
- Levegő hőmérséklete
- Levélnedvesség
A levegő hőmérséklete mellett a levélnedvesség is irányadó tényező lehet, mivel a hosszú levélnedvességi időszakok antagonista gomba (Ampelomyces quisqualis) megtelepedéséhez vezetnek, ami az U. necator csökkenéséhez vezet. A modell alapjai megegyeznek a kaliforniai kockázati modellel, de ebben a modellben a levélnedvességet is figyelembe veszik. A 8 óránál hosszabb ideig tartó levélnedvesség 10 pont csökkenést eredményez.
Az alacsony, 0~20-as indexértékek azt jelzik, hogy a kórokozó nem szaporodik. A 20~60-as index közepesnek tekinthető, és a szokásos permetezési időköz érvényes. A 60 feletti indexértékek azt jelzik, hogy a kórokozó gyorsan (5 naponta) szaporodik, és nagy a járványveszély, ezért a permetezési időköz lerövidítése ajánlott.
Irodalom
- Gadoury, D. M., & Pearson, R. C. (1990). Aszkospórák dehiszenciája és aszkospórakibocsátás az Uncinula necatorban. Phytopathology, 80(4), 393-401.
- Gadoury, D. M., & Pearson, R. C. (1990). Az aszkospórák csírázása és a Vitis fertőzése az Uncinula necator által. Phytopathology, 80(11), 1198-1203.
- Hall, T. W. (2000). A szőlő lisztharmat, Uncinula necator, epidemiológiája a Willamette-völgyben.
- Halleen, F., & Holz, G. (2001). Az Uncinula Necator (lisztharmat) biológiájának, epidemiológiájának és védekezésének áttekintése a szőlőn, tekintettel Dél-Afrikára. South african journal of Enology and Viticulture, 22(2), 111-121.
- Rügner, A., Rumbolz, J., Huber, B., Bleyer, G., Gisi, U., Kassemeyer, H. H., & Guggenheim, R. (2002). Az Uncinula necator telelőszerkezetek kialakulása és a szőlő kolonizációja szabadföldi körülmények között. Növénykórtan, 51(3), 322-330.
Kórokozó
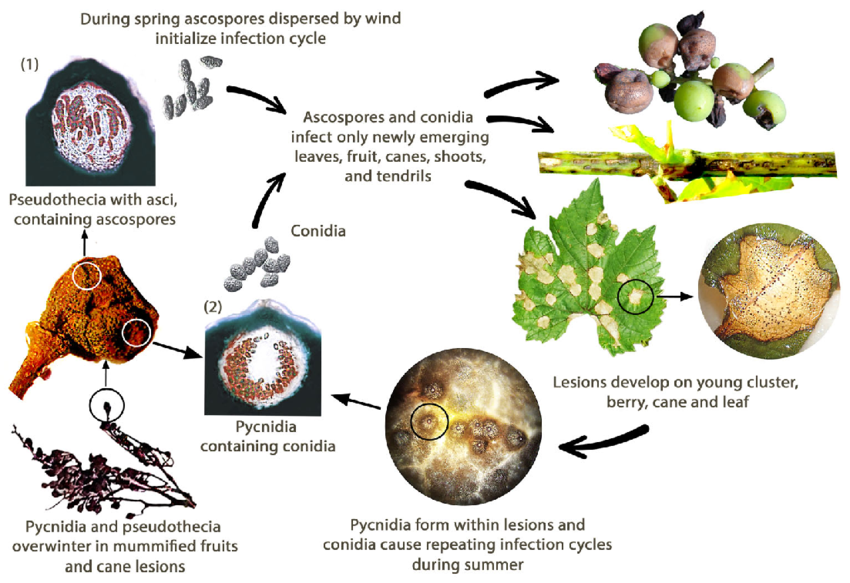
A szőlő fekete rothadását a Guignardia bidwellii gomba okozza. A gomba a szőlő különböző részein telel át, és a fertőzött hajtások sérüléseiben több mint két évig is életben maradhat.
Tavasszal a piknídiumok konídiumokat (ivartalan spórákat), az álgubók pedig aszkospórákat (ivaros spórákat) termelnek. A konídiumok jellemzően rövid távolságra terjednek, míg az aszkospórák nagyobb távolságra is fertőzhetnek. Ezeket a spórákat a szél és az eső szállítja, hogy megfertőzzék a fogékony gazdaszervezetek fiatal szöveteit, és primer fertőzéseket indítsanak el. A konídiumok másodlagos inokulumként is szolgálnak, hozzájárulva a betegség gyors és ismételt terjedéséhez.
Augusztus folyamán a piknídiumok átváltoznak egy telelő stádiumba, amely pszeudotéciumokat termel, amelyek aszkospórákat hoznak létre. Ezek az aszkospórák fontos forrásai a következő tavaszi elsődleges fertőzéseknek.
Tünetek

Vörösesbarna foltok alakulnak ki, amelyek végül összeolvadnak, gyakran apró fekete pontokkal körülvéve, amelyek a konídiumokat termelő, piknídiumoknak nevezett gombaszerkezetek. A fiatal és gyorsan növekvő levelek különösen fogékonyak erre a fertőzésre.
A gyümölcsfertőzés jellemzően a kehely lehullása után jelentkezik, és a legtöbb tünet akkor jelentkezik, amikor a gyümölcs már félig vagy majdnem teljesen kifejlődött. Kezdetben apró foltok jelennek meg, amelyeket barna gyűrű vesz körül, majd a betegség előrehaladtával megnőnek és elsötétednek, és végül az egész bogyót beborítják. Néhány napon belül a fertőzött bogyók mumifikálódhatnak, összetörhetnek vagy idő előtt lehullhatnak.
Ezenkívül más növényi részek, például a hajtások, a szárak és az indák is érintettek. Ovális alakú, a lilától a feketéig változó színű sérülések alakulnak ki, és a piknídiumok elszóródnak a sérülések felületén.
FieldClimate modell
Szőlészet Fekete rothadás modell
Szükséges érzékelők:
- Levegő hőmérséklete
- Relatív páratartalom
- Levélnedvesség
- Csapadék
A modell eredetileg a Spotts által publikált szakirodalmon alapul, de Daniel Molitor módosításával. Az átdolgozás három súlyossági osztályt (könnyű, közepes és súlyos) vezetett be, hasonlóan az almarothadás modelljeinek kialakításához. Meleg hőmérsékleten a 8 órás levélnedvesedési időszak elegendő a fertőzéseknek való kedvezéshez, és a fertőzést befejezettnek tekintik, amikor egy grafikon eléri az 100% értéket.
A fertőzés súlyossága a gomba optimális hőmérsékletének és nedvességtartalmának időtartamától függ. A Spotts-kritériumoknak megfelelő fertőzéseket könnyű fertőzésnek minősítik. A 150% és a 200% által a Spott-kritériumoknak megfelelő fertőzéseket közepes, illetve súlyos fertőzésnek minősítik.
A fertőzési nyomástól függően a fertőzéseket megelőzően kell lefedni, vagy röviddel a fertőzés után gyógyító permetezést kell alkalmazni. A Mosel vagy a legtöbb osztrák szőlőtermő terület mérsékelt, félszáraz éghajlatán nem ajánlott a permetezés az enyhe fertőzés megállapítása után.
Irodalom
- Wilcox, Wayne F. "Fekete rothadás Guignardia bidwellii". 102GFSG-D4 számú betegségazonosító lap. 2003. Cornell. 2010. október 24.
- http://www.nysipm.cornell.edu/factsheets/grapes/diseases/grape_br.pdf
- Ellis, Michael A. "Tájékoztató a mezőgazdasági és természeti erőforrásokról: Grape Black Rot." Növénykórtani Tanszék. Az Ohiói Állami Egyetem Állami Hagyományőrző Központja. 2008
- http://ohioline.osu.edu/hyg-fact/3000/pdf/HYG_3004_08.pdf
- Molitor, D. (2009). Untersuchungen zur Biologie und Bekämpfung der Schwarzfäule (Guignardia bidwellii) an Weinreben. Gesellschaft zur Förderung der Forschungsanstalt Geisenheim.
- Ries, S. M. (1999). Növénybetegségekről szóló jelentések: A szőlő fekete rothadása. Integrated Pest Management at the Uni. of Illinois. http://ipm. illinois. edu/diseases/series700/rpd703.
- Spotts, R. A. (1977). A levélnedvesség időtartamának és a hőmérsékletnek a hatása a Guignardia bidwellii fertőzőképességére a szőlőleveleken. Phytopathology, 67(11), 1378-1381.
- Szabó, M., Csikász-Krizsics, A., Dula, T., Farkas, E., Roznik, D., Kozma, P., & Deák, T. (2023). A szőlő fekete rothadása (Guignardia bidwellii) - Átfogó áttekintés. Horticulturae, 9(2), 130.
- http://extension.cropsciences.illinois.edu/fruitveg/pdfs/771-BlackRotOfGrape.pdf
- https://www.missouribotanicalgarden.org/gardens-gardening/your-garden/help-for-the-home-gardener/advice-tips-resources/insects-pests-and-problems/diseases/fruit-spots/black-rot-of-grapes
Kórokozó

Lobesia botrana a régiótól függően általában évente két-három generáción keresztül fejlődik. Az első nemzedék (május és június) csak a virágokat érinti, ezért a kezelés csak akkor szükséges, ha a molypopuláció különösen magas. A második (júliusi és augusztusi) és a harmadik generáció okozza a legnagyobb kárt, a harmadik generáció különösen káros, mivel egybeesik a szőlő érésével. A negyedik nemzedék melegebb éghajlaton előfordulhat, de a kezelés általában nem szükséges, mivel az a szőlőszürethez igazodik.
A lepke bábként telel át a gubókban, és amikor a hőmérséklet tavasszal emelkedik, az első generáció felnőtt példányai kelnek ki, a hímek általában előbb jelennek meg, mint a nőstények. Az első nemzedék repülése a rügyfakadás közelében kezdődik és 4-5 hétig tart, ezalatt történik a párzás. A párosodás után egy-két nappal a nőstények 80-160 tojást raknak.
A 0,6-0,8 mm átmérőjű tojások szabad szemmel is láthatóak. Kezdetben krémfehérek, majd az embrió fejlődésével sárgává válnak, a lárva fejének színe fekete. A tojások 66 Celsius-fokos nap (DDC) után kelnek ki, körülbelül 7-11 nap múlva. A lárvák együtt hálózzák be a virágrészeket, fejlődésük 20-30 napig tart. Ezután következik a bábozódás, és a kifejlett egyedek 6-14 nappal később kelnek ki.
A "foknapot" úgy számítják ki, hogy mérik, hogy a napi átlaghőmérséklet mennyivel haladja meg a fejlődéshez szükséges bizonyos alapküszöbhőmérsékletet. Ahogy a hőmérséklet az alapérték fölé emelkedik, úgy halmozódnak a foknapok.
A kifejlett lepkék 6-8 mm hosszúak, szárnyfesztávolságuk 11-13 mm. A nőstények valamivel nagyobbak, mint a hímek, bár mindkettőnek hasonló mozaikos mintázatú szárnya van.
A második és harmadik generáció tojásai gyorsabban, 3-5 napon belül kelnek ki. A nőstények egyenként rakják le a tojásokat az árnyékos bogyókra, és a lárvák a kikelés után behatolnak a bogyókba, és kivájják azokat. Ősszel a 11 óránál hosszabb éjszakák diapauzát váltanak ki, egy olyan nyugalmi állapotot, amely lehetővé teszi a bábok számára, hogy jobban elviseljék a hideget, és így át tudjanak telelni.
Tünetek
Az első generáció hernyói a szőlő virágzatán táplálkoznak, a virágbimbókat kivájják és védekezésképpen megfonnyasztják, ami a fürtön belüli károsodáshoz vezet. A második generációban a lárvák a fejlődő bogyókat veszik célba, amelyeken a gombakórokozók, mint például a Botrytis cinerea. Ez a közvetlen táplálkozás látható károkat okoz, amelyeket jellemzően sötét foltok jeleznek a táplálkozási helyek körül. A harmadik nemzedék okozza a legjelentősebb kárt, mivel a lárvák behatolnak az érő bogyókba, és ott táplálkoznak.
Megszáradt bogyók és lárvaürülék marad. A bogyó epidermális szövetének foszlányai lazán a bogyószárhoz rögzülve maradnak, a száraz, üreges bogyó "bőrével" együtt.
FieldClimate modell
Grapevine Berry Moth modell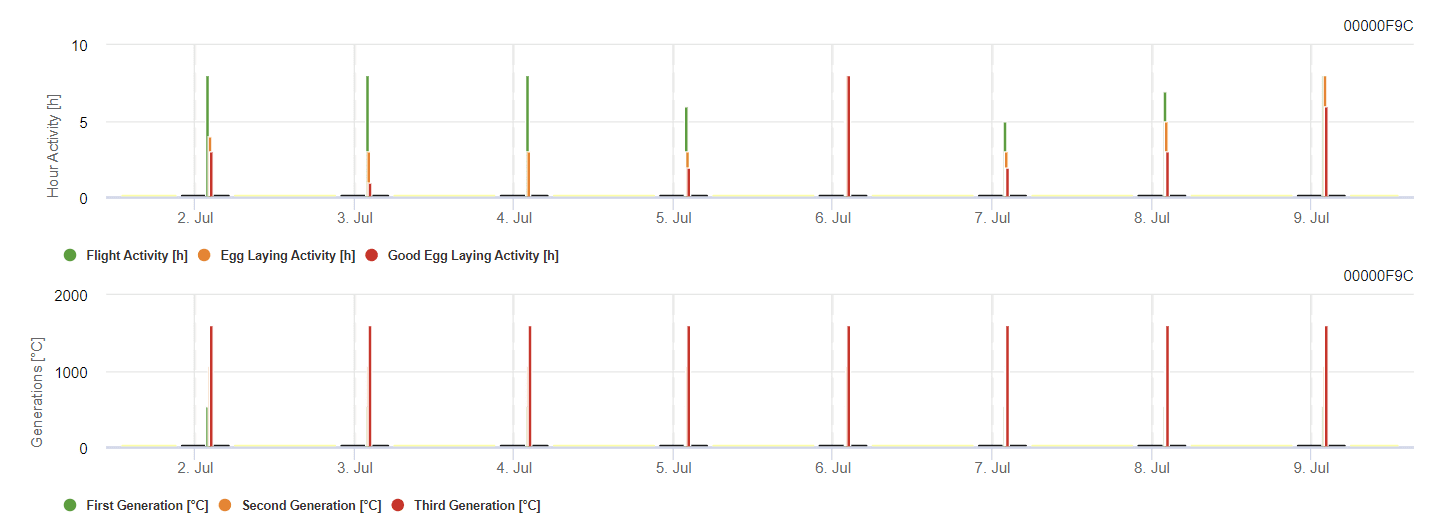
Szükséges érzékelő:
- Levegő hőmérséklete
A modell a kockázatot a levegő hőmérséklete alapján számítja ki. Az első grafikon a rovar számára a tojástermeléssel és a jó tojástermelési aktivitással járó repülési aktivitási időszakokat mutatja. A második grafikon azt mutatja, hogy az adott időszakban melyik generációra kell számítani. A harmadik grafikonon az éghajlati adatok jelennek meg, amelyek a foknapok halmozódását mutatják (8 °C-on felüli hőmérséklet 24 °C-ig óránként osztva 24-gyel).
Irodalom
- Varela, L. G., Smith, R. J., Cooper, M. L., & Hoenisch, R. W. (2010). Az európai szőlőmoly, Lobesia botrana. Napa-völgyi szőlőültetvények. Pract. Winery Vineyard, 2010, 1-5.
- https://ipm.ucanr.edu/invasive-and-exotic-pests/european-grapevine-moth/
- https://www.ages.at/en/plant/plant-health/pests-from-a-to-z/grape-berry-moth
Kórokozó
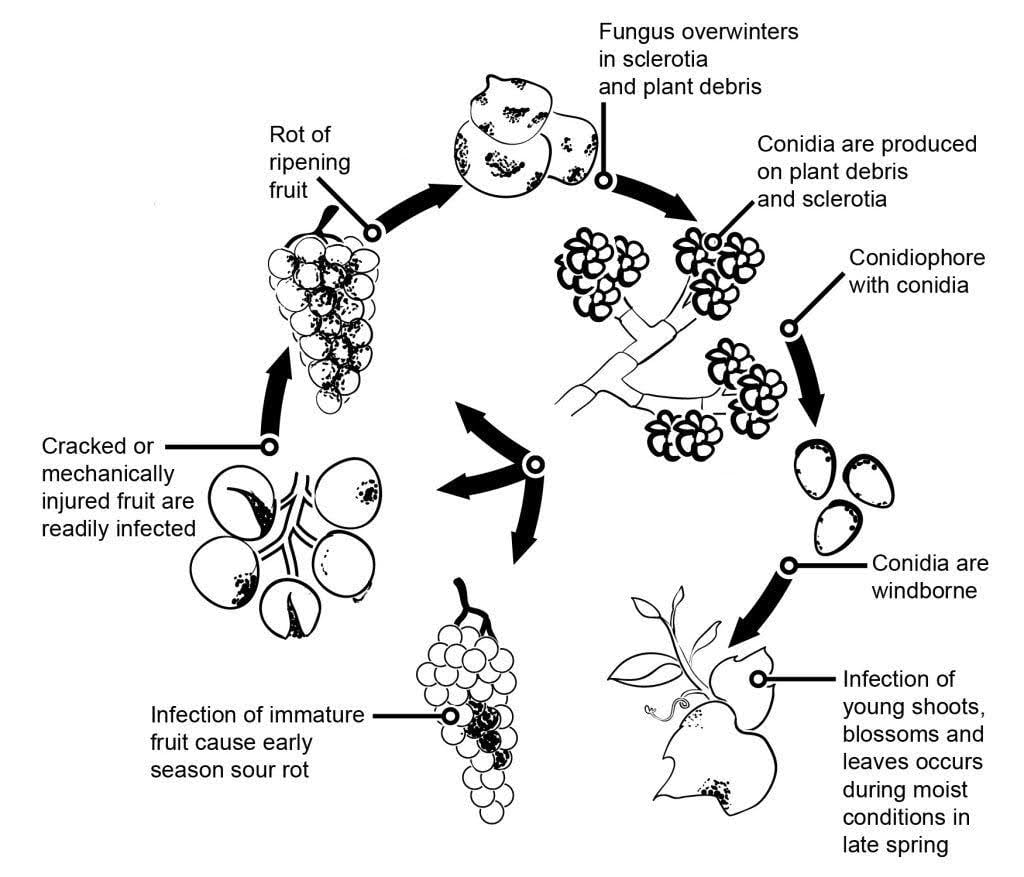
A szürkepenész kórokozója a szőlőben a következő Botrytis cinerea.
A szürkepenész járványtanát jelentősen befolyásolják a virágfertőzések által kezdeményezett látens fertőzések. Különböző virágtól a gyümölcsig terjedő fertőzési utakat azonosítottak: A gomba megfertőzi a virág porzóját, és eléri a petecsomót, ahol a növény előre kialakított védekező mechanizmusa miatt látens marad. A porzókon, szirmokon és sziromleveleken keresztül történő fertőzés szintén kritikus. A B. cinerea képes megfertőzni a porzókat, és szisztematikusan növekszik a termőedény felé, átterjedve a bogyókban a virágszárra és az érzőszövetekre. A kórokozó szaprofitikus növekedése más útvonalakon is előfordul.
Tavasszal a telelő micéliumok vagy szkleróciumok a fertőzés elsődleges forrásai. A konídiumok (ivartalan spórák) a szél és az eső által termelődnek és terjednek. Leszálláskor a konídiumok kicsíráznak, csíracsöveket és appressóriumokat képezve, amelyek behatolnak a növénybe és megfertőzik azt. A gomba látens maradhat, amíg a gyümölcs be nem érik és a cukortartalom meg nem nő. Az elsődleges inokulumforrásokból származó konídiumok a beindulás, a termelés és a terjedés napszakos ciklusát követik.
Míg a B. cinerea ivaros stádiuma ritkán figyelhető meg, a kórokozó a bogyók érésével egyre aktívabbá válik. A bogyók megnövekedett cukortartalma növeli a fertőzésre való fogékonyságot. Ugyanígy a virágok is nagyon fogékonyak, amikor elszáradnak, a bőséges pollen pedig növeli a fertőzés súlyosságát.
Tünetek

A szürkepenész elsősorban az érett szőlőbogyókat fertőzi meg, amelyek kezdetben puhának és vízzel átitatottnak tűnnek. Idővel a bogyók vörösesbarnára színeződnek és összezsugorodnak. Kedvező körülmények között a bogyókat szürke gombamirigy- és konídiumtömegek borítják. Az egészséges bogyók a beteg bogyókkal való közvetlen érintkezés révén is megfertőződhetnek.
A gomba virágfoltosságot is okozhat, ami a szezon elején jelentős terméskiesést okozhat. Bár a levélfertőzések ritkák, ha mégis előfordulnak, az erek mentén tompa zöld foltokként indulnak, amelyek végül nekrotikus elváltozásokká alakulnak.
FieldClimate modell
Általános szürke szerszám modell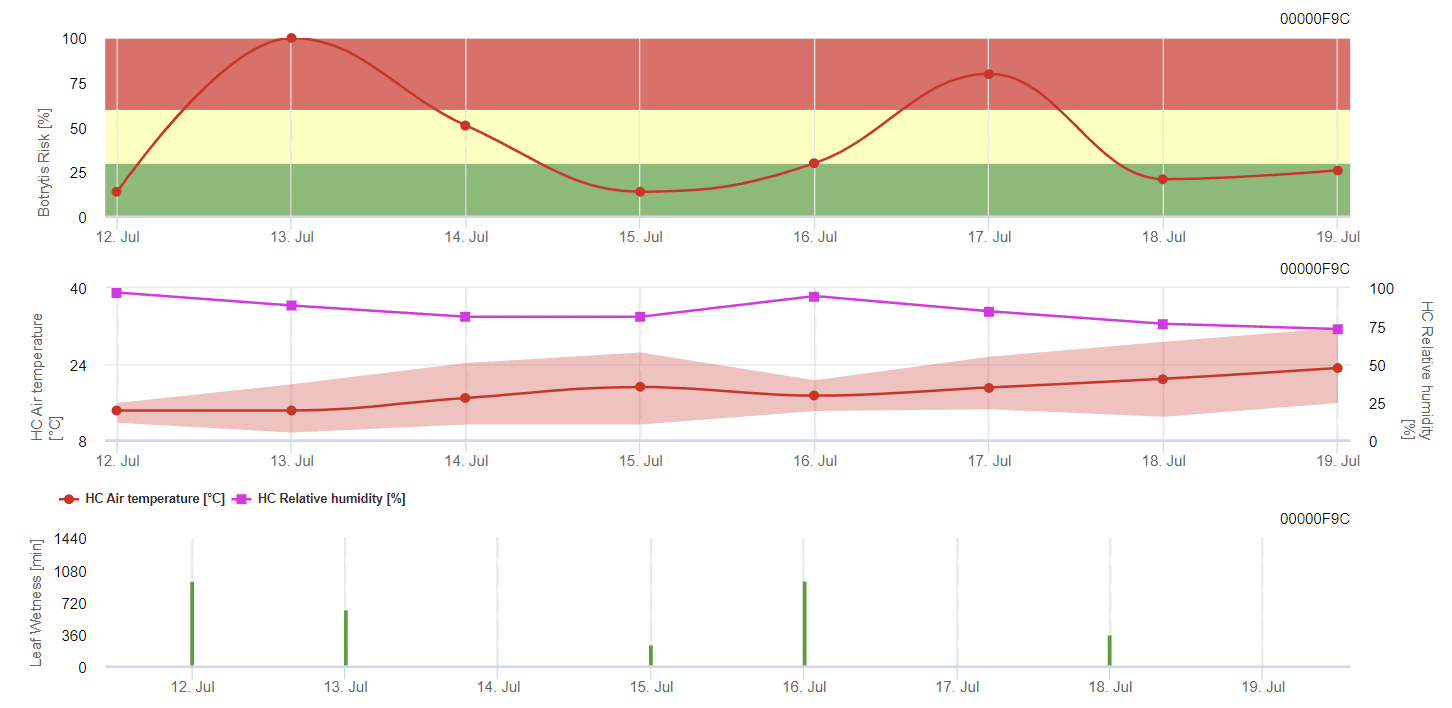
Szükséges érzékelők:
- Levegő hőmérséklete
- Relatív páratartalom
- Levélnedvesség
B. cinerea a nedves éghajlathoz kapcsolódik. A gombának magas relatív páratartalomra vagy szabad víz jelenlétére van szüksége a fertőzéshez. A hosszabb levélnedvességgel járó meleg időszakok a B. cinerea kockázatának növekedéséhez vezetnek, míg a száraz időszakok csökkentik azt. A fertőzés a fiatal hajtásokon a hosszú nedves időszakok vagy a káros jégesők idején történik.
A modell a kockázatot 0 és 100% közötti értékben számítja ki. Ez az érték jelzi a nyomást B. cinerea az adott időpontban - ha 100% értéket kapunk, az azt jelenti, hogy többször volt elég hosszú nedves időszak ahhoz, hogy a fogékony szöveteket megfertőzze. A levélnedvesség és a hőmérséklet között úgynevezett "nedves pontokat" számolunk, amelyek maximuma kezdetben 38400 pont (a szezon kezdete, amely 30% kockázatot mutat). Ezt követően minden egyes, körülbelül 4000 nedves pontot tartalmazó nedves időszak 10%-vel növeli a kockázatot, illetve a másik oldalon minden egyes száraz időszak a korábbi érték ⅕-jával csökkenti a kockázatot.
Irodalom
- Broome, J. C., English, J. T., Marois, J. J., Latorre, B. A., & Aviles, J. C. (1995). A szőlő Botrytis fürtrothadásának fertőzési modelljének kidolgozása a nedvességtartam és a hőmérséklet alapján. Phytopathology, 85(1), 97-102.
- Elmer, P. A., & Michailides, T. J. (2007). A Botrytis cinerea epidemiológiája gyümölcsösökben és szőlőkben. In Botrytis: biológia, patológia és védekezés (pp. 243-272). Dordrecht: Springer Netherlands.
- Williamson, B., Tudzynski, B., Tudzynski, P., & Van Kan, J. A. (2007). Botrytis cinerea: a szürkepenész-betegség oka. Molecular plant pathology, 8(5), 561-580.
- Ciliberti, N., Fermaud, M., Roudet, J., & Rossi, V. (2015). A környezeti feltételek jobban befolyásolják az érett szőlőbogyók Botrytis cinerea fertőzését, mint a törzs vagy a transzpozon genotípus. Phytopathology, 105(8), 1090-1096.
Kórokozó
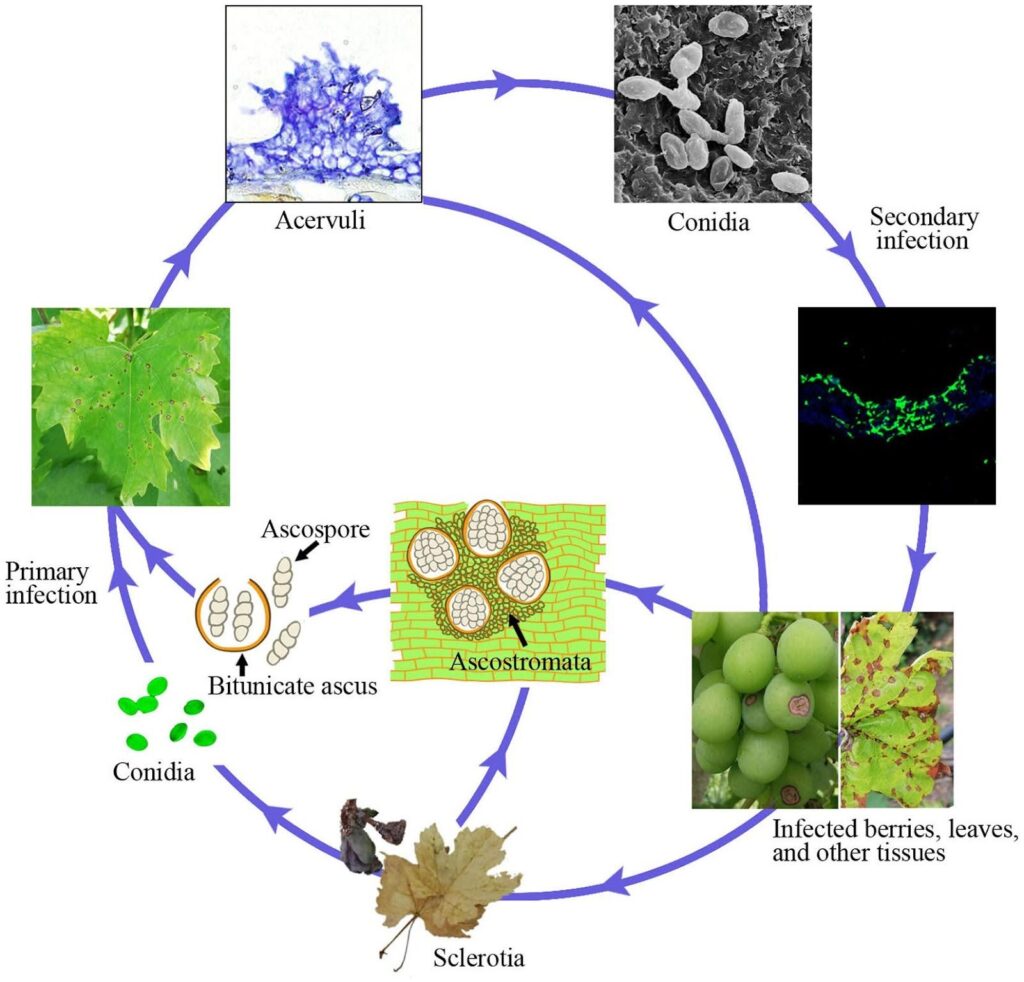
Az antraknózist a szőlőben a kórokozó okozza. Elsinoe ampelina.
A betegség fő forrása a fertőzött vesszők. A télen a sérülésekben és a bogyókban túlélő szkleróciumok és micéliumok tavasszal aktívvá válnak, és nedves körülmények között (eső vagy 24 órán át tartó harmat) 2~40°C hőmérséklet-tartományban aszkospórákat (ivaros spórák) és konídiumokat (ivartalan spórák) termelnek.
Ezek a spórák az eső és a szél által új szövetekbe szóródnak, és a leszállás után kicsíráznak, csírákat és appressóriumokat képeznek, és új fertőzést indítanak el. Megfertőzhetik az új leveleket, hajtásokat, indákat és fiatal bogyókat. A meleg időjárás csökkenti a kezdeti fertőzéshez és a lappangási időszakhoz szükséges nedves időtartamot. A spórák optimális csírázása 25~30°C-on következik be, és legalább 3~4 óra levélnedvesség szükséges. A 10°C körüli hőmérsékleten történő fertőzéseknél a lappangási idő körülbelül 14 nap.
A kolonizáció előrehaladtával akervulusok jelennek meg és új konídiumok termelődnek, amelyek másodlagos fertőzési inokulumként szolgálnak. Ezek felelősek a gyors és további fertőzésért a szezonban.
Tünetek
E. ampelina a szőlő lombos, zamatos részeit támadja meg, beleértve a hajtásokat, leveleket, levélnyél, indák, indák, vesszők és bogyók; a leggyakoribbak a hajtások és a bogyók sérülései. A szőlő ontogén rezisztenciával rendelkezik a gombával szemben, így a fiatal szövetek fogékonyabbak.
Először kis vörösesbarna foltok jelennek meg, amelyek nedves időben megnagyobbodnak, és kissé beesnek, szürke középponttal, amelyet sötét szegély vesz körül. Néha a tünetek jégeső okozta sérülésnek tűnhetnek. A levelek nekrotikus közepe általában kiesik, ami "lövészlyuk" megjelenést eredményez.
Súlyos fertőzés esetén a növények korán lombtalanodhatnak, a vessző növekedése lelassulhat, a szár törhet, a bogyók lehullhatnak, és a bogyók fejlődése és érése késhet.
FieldClimate modell
Szőlészeti antraknózis modell
Szükséges érzékelők:
- Levegő hőmérséklete
- Relatív páratartalom
- Levélnedvesség
- Csapadék
A modell a levegő hőmérsékletének, a relatív páratartalomnak, a levélnedvességnek és a csapadéknak a figyelembevételével számítja ki az antraknózis kockázatát.
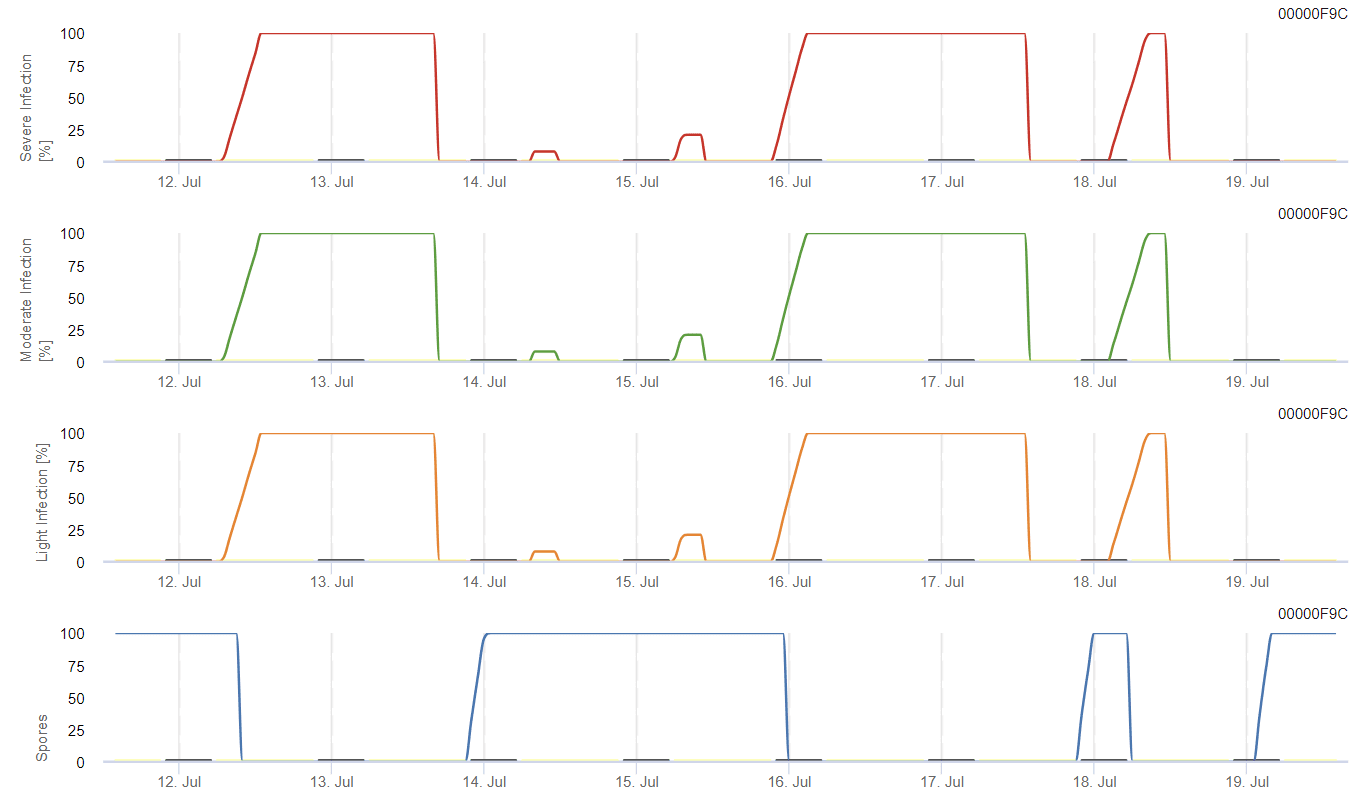
A gyenge, közepes és súlyos fertőzés lehetséges kialakulását külön grafikonok mutatják. A fertőzést akkor kell teljesnek tekinteni, amikor a növekvő görbe eléri az 100% értéket. Az utolsó grafikon a sporulációt mutatja E. ampelina; ha az ottani érték eléri az 100% értéket, akkor feltételezzük, hogy a spórák a E. ampelina jelen vannak.
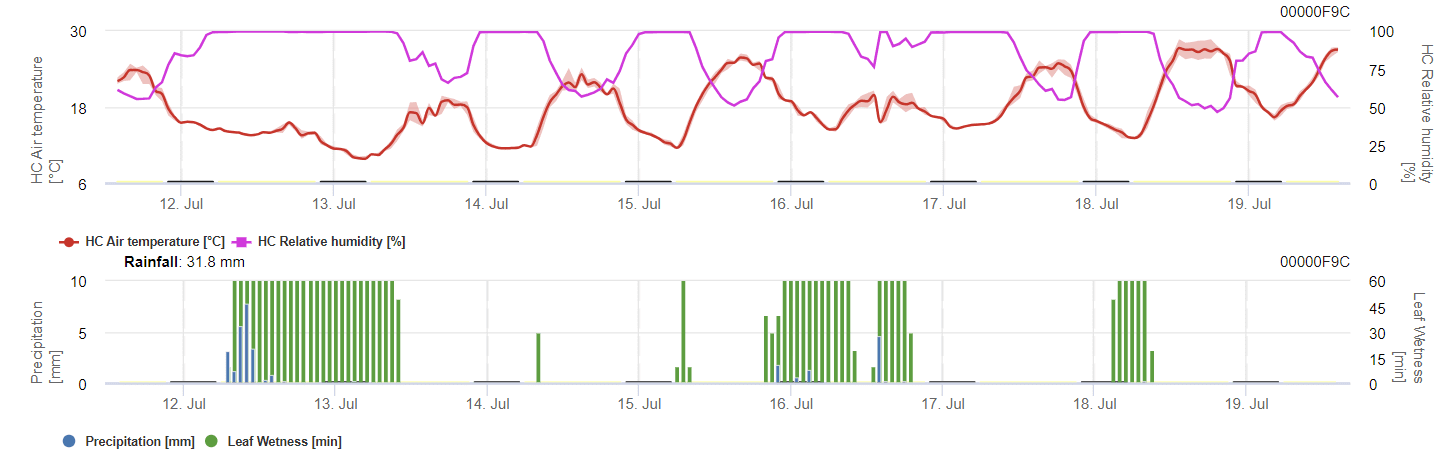
Az áttelelő spórák fejlődéséhez a körülményeknek meg kell felelniük a 2 és 40°C közötti hőmérséklet-tartománynak, a 90% feletti relatív páratartalomnak vagy a levélnedvességnek. Amint a spórák fejlődése eléri az 100% értéket, a fertőzés számítása megkezdődik. A fertőzés súlyossága a nedves körülményektől (esőzések) függ. Ha azonban a páratartalom 50% alá csökken, mind a spóraképződés, mind a fertőzés számítása leáll.
Irodalom
- Li, Z., Dos Santos, R. F., Gao, L., Chang, P., & Wang, X. (2021). Az Elsinoe ampelina által okozott szőlőantraknózis jelenlegi helyzete és jövőbeli kilátásai: Fontos betegség a nedves szőlőtermő régiókban. Molecular Plant Pathology, 22(8), 899-910.
Kórokozó
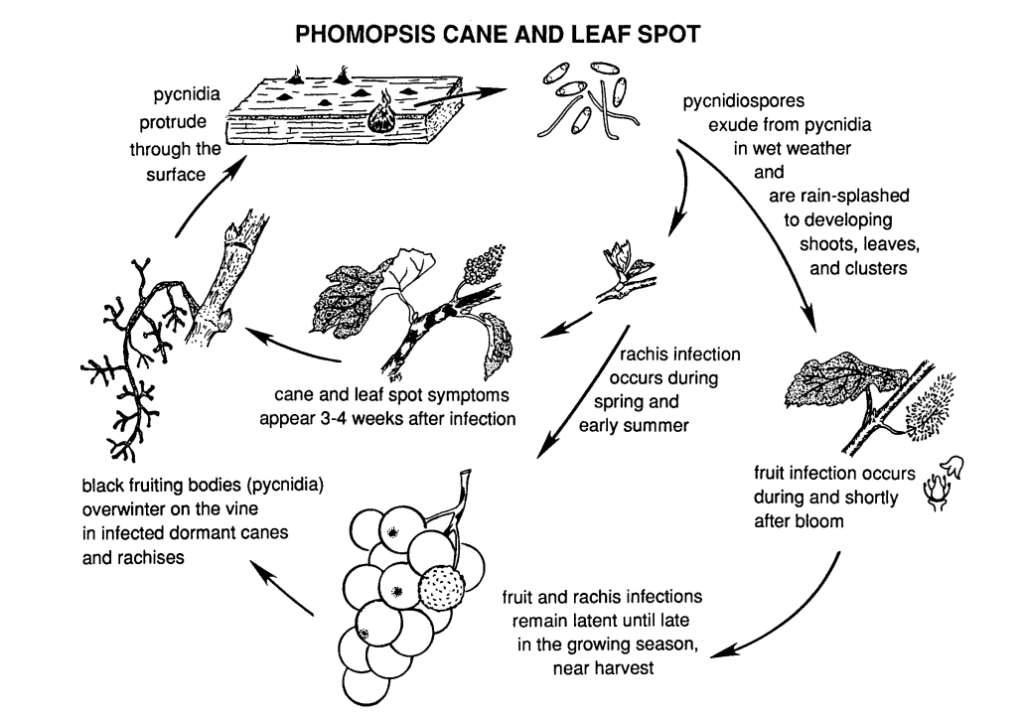
A Phomopsis vessző- és levélfoltosságot a szőlőben a következők okozzák Phomopsis viticola.
A gomba a szőlő fás részein telel át, és a következő tavasszal újra aktívvá válik. Amint a hőmérséklet emelkedik és a tavaszi esőzések megkezdődnek, a spórák a téli átvészelő szerkezetekből, az úgynevezett piknídiumokból szabadulnak fel. Ahogy a spórák az eső és a szél révén új szövetekre kerülnek, a fertőzés terjed.
Az elsődleges fertőzési időszak jellemzően a kora tavaszi esőzésekkel esik egybe, és röviddel a rügyfakadás után kezdődik, amikor a hajtások a növekedés korai szakaszában vannak. A fiatal hajtások, a hajtáscsúcsok, a termésfürtök és a vessző mind érzékenyek a fertőzésre. Bár a hajtáscsúcsok az egész vegetációs időszak alatt fertőződhetnek, a fertőzések a rügyfakadástól a virágzásig a leggyakoribbak. A gyümölcsökben a kórokozó a gyümölcsök éréséig nyugalmi állapotban maradhat. A lappangási idő általában 3-4 hétig tart.
Tünetek
A leveleken és levélnyélen apró, fekete közepű, sárga szegéllyel körülvett sötét foltok jelennek meg, amelyek végül összeolvadnak. Az alaplevelek eltorzulhatnak és gyűrötté válhatnak, és előfordulhat, hogy nem fejlődnek ki teljes méretükre. Ha a levélnyél erősen fertőzött, megsárgul és lehullik, ami levélhulláshoz vezet. A levelek ernyőhatást keltenek, ami csökkenti a spórák további fertőzéseit.
Hasonló foltok jelennek meg a hajtásokon, ami a fertőzés helyén a hámrétegek repedezését okozza. Ahogy a foltok összeolvadnak, a fertőzött hajtásrészek sebhelyes szerkezetűvé válnak. Súlyos esetekben a hajtások satnyulhatnak, letörhetnek vagy akár el is pusztulhatnak.
A fonák sérülései miatt a fonák megsüllyed és törékennyé válik. A nyári nyugalmi időszakot követően a gomba kora ősszel újra aktivizálódik, ami bogyó- és fürtrothadáshoz vezet. Bár a gyümölcsök általában nem fertőződnek nagymértékben, a bogyókon foltok alakulnak ki, amelyeket gyakran fekete foltok (piknídiumok) kísérnek a bogyóhéjon. Nedves körülmények között sárga spóratömegek szivároghatnak ki a piknídiumokból, ami a gyümölcs elszíneződéséhez, érdes szerkezetéhez és mumifikálódásához vezet.
A fertőzött fa a nyugalmi időszakban kifehéredettnek tűnik. A súlyosan fertőzött vesszők és hajtások sötét elszíneződéseket mutatnak a kéreg fehérített területeivel vegyesen. A pycinidiumok a felszínen is áttörhetnek. Az alacsony hőmérséklet, valamint a károsodott floem- és xilémfunkció meggyengíti és elpusztítja a vesszőket, a hajtásokat és a rügyeket.
FieldClimate modell
Szőlészeti fomopszis fertőzési modell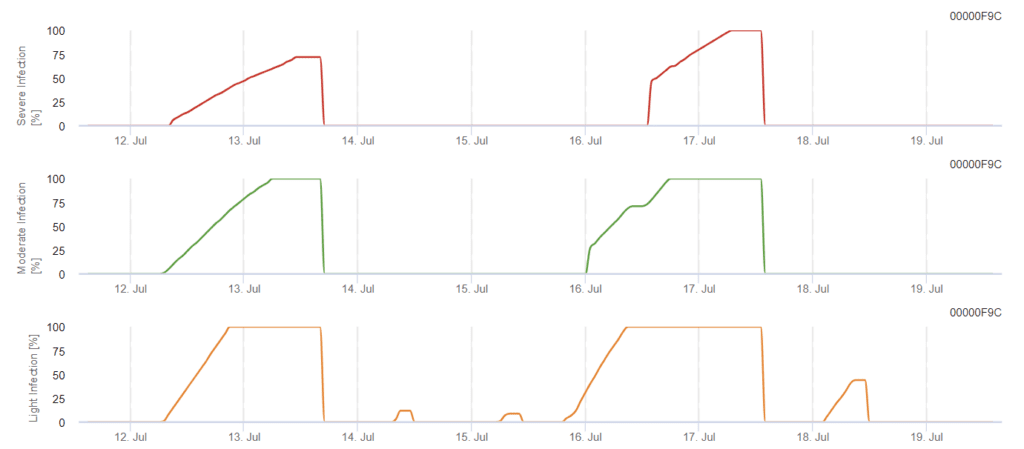
Szükséges érzékelők:
- Levegő hőmérséklete
- Relatív páratartalom
- Levélnedvesség
- Csapadék
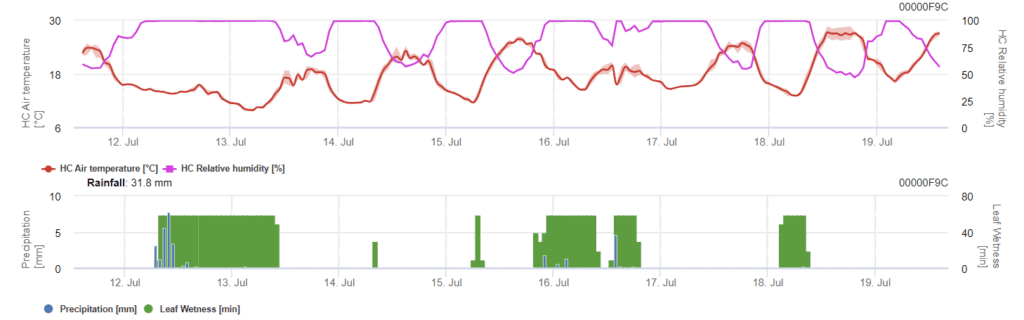
A modell a Phomopsis-fertőzés kockázatát 5 és 35 °C közötti hőmérséklet-tartományban és nedves körülmények között (levélnedvesség, magas relatív páratartalom) határozza meg. A fertőzés súlyossága a csapadék mennyiségétől (2 mm-nél több) függ, mivel a spórák gyorsabban terjednek az egészséges növényi anyagban. Az 100% fertőzés kimutatása azt jelzi, hogy a szántóföldön optimális feltételeket mértek a növényi szövetek megfertőzéséhez, ezért a növényvédelmi méréseket figyelembe kell venni.
Irodalom
- https://agriculture.vic.gov.au/biosecurity/plant-diseases/grapevine-diseases/phomopsis-cane-and-leaf-spot-of-grapevines
- Bettiga, L. J. (szerk.). (2013). Szőlő kártevők elleni védekezés (3343. kötet). UCANR Publications.
- Pscheidt, J. W., & Pearson, R. C. (1991). Phomopsis vessző- és levélfoltosság.