Rez sójová, konkrétně asijská rez sójová, je závažné onemocnění sóji způsobené houbovým patogenem. Phakopsora pachyrhizi. Rozšíření onemocnění: Rozšíření: Asie a Austrálie, nověji Afrika, Severní Amerika a Jižní Amerika. Nemoc byla poprvé pozorována v Japonsku v roce 1902. Přes Ameriku se patogen rozšířil po celé Asii a Austrálii. Rez sójová nemůže přezimovat v oblastech s mrazivými teplotami, ale může se rychle šířit větrem na tak velké vzdálenosti, její vývoj může být tak explozivní a může způsobit tak rychlou ztrátu listů, že je nyní jednou z nejobávanějších chorob v oblastech pěstování sóji na světě.
Příznaky a znaky
První příznaky rzi sójové způsobené Phakopsora pachyrhizi začínají jako velmi malé hnědé nebo cihlově červené skvrny na listech. Na poli se tyto skvrny obvykle objevují v dolní části koruny v době květu nebo po něm, i když za určitých okolností mohou být infikovány i sazenice. První léze se často objevují u báze listu v blízkosti řapíku a listových žilek. Tato část listu pravděpodobně déle zadržuje rosu, což vytváří příznivější podmínky pro infekci. Léze zůstávají malé (2-5 mm v průměru), ale s postupující chorobou se jejich počet zvětšuje. V těchto lézích, většinou na spodní straně listu, se tvoří puchýřky, zvané uredinia, které mohou produkovat mnoho urediniových spor.
Vyvýšené puchýřky jsou viditelné pouhým okem, zejména při sporulaci. I když jsou léze malé, každá léze má často několik pustul (uredinií) . Léze mohou být zcela pokryty urediniovými sporami, když jsou pustuly aktivní. Urediniospóry rzi sójové jsou světle žlutohnědé až bezbarvé, s echinulární (krátkými ostny) povrchovou ornamentikou. Toto zbarvení se liší od mnoha jiných patogenů rzi, jejichž spory jsou často červenohnědé (rezavě zbarvené). Klíčení P. pachyrhizi urediniospory pronikají rovníkovým (centrálním) pórem a vytvářejí zárodečnou trubičku, která je zakončena apresoriem, jímž houba proniká do hostitele přímo nebo stomií.
Jak se na listu tvoří stále více a více lézí, postižené místo začíná žloutnout a nakonec list z rostliny odpadne. Zatímco rez sóji obvykle začíná v dolní části koruny, rychle postupuje vzhůru po rostlině, dokud nejsou všechny listy do určité míry postiženy chorobou. Silně napadené rostliny mohou být zcela olistěné. Ztráta účinných listových pletiv má za následek snížení výnosu v důsledku menšího počtu semen i menšího množství osiva. Byly zaznamenány ztráty výnosu až 30 až 80%, ale výše ztrát závisí na tom, kdy choroba začíná a jak rychle postupuje. Kromě listů se rez sóji může objevit také na řapících, stoncích a dokonce i na listenech, ale většina rzí se vyskytuje na listech.
Léze mohou být opálené nebo červenohnědé. Opálené léze mají mnoho pustul, které produkují četné urediniospóry. Červenohnědé léze, které jsou považovány za reakci mírné rezistence, mají pouze několik pustul, které produkují pouze několik urediniospór. Jak bude popsáno v části Management chorob, tento typ lézí závisí na kmeni patogenu a může se objevit na stejném listu jako opálené léze nebo se opálené léze mohou s věkem změnit na červenohnědé. Symptomy a příznaky na jiných hostitelích, jako je kudzu, jsou podobné, i když velikost lézí se může lišit. Stárnutím mohou puchýřky zčernat. To je způsobeno tvorbou vrstvy teliospor v puchýřcích, čímž se puchýřky mění z uredinií na telia.
Teliospory mají dvě funkce: přežití houby v nepřítomnosti živého hostitele (overseasoning) a pohlavní rozmnožování. Silné stěny teliospor chrání houbu před okolním prostředím a útoky jiných organismů. U rzí teliospory klíčí a vytvářejí bazidium a čtyři bazidiospory, během nichž dochází k pohlavní rekombinaci. Klíčení P. pachyrhizi Teliospory byly pozorovány pouze v laboratoři a nezdá se, že by významně přispívaly k šíření této choroby v terénu.
Biologie patogenů
Existují dvě blízce příbuzné houby, které způsobují rez na sóji: Phakopsora pachyrhizi, někdy označovaný jako asijský nebo australský patogen rzi sójové, který se však nyní vyskytuje i na západní polokouli, a P. meibomiae, takzvaného patogenu rzi sóji Nového světa, který se vyskytuje pouze na západní polokouli. Až na několik drobných znaků se obě houby zdají být morfologicky identické, ale P. pachyrhizi je mnohem agresivnější na sóju než na boby. P. meibomiae. K dnešnímu dni, P. meibomiae nebylo prokázáno, že by ve Střední a Jižní Americe způsobovala významné ztráty na výnosech. Oba druhy lze rozlišit pomocí protokolů analýzy DNA.
Stejně jako ostatní rzi jsou i patogeny rzi sójové obligátními parazity, kteří ke svému růstu a rozmnožování potřebují živého hostitele. V přírodních podmínkách mohou přežívat mimo hostitele jako urediniospory pouze několik dní. Oba patogeny rzi sójové podle našich nejlepších znalostí produkují pouze dva typy spor: urediniospory a teliospory (obrázek 15). To je v kontrastu s jinými rzí, které mohou mít až pět stádií spor (například rez stonková pšenice). U rzi sójové, stejně jako u většiny rzí, je urediniové stadium opakujícím se stadiem. To znamená, že urediniové spory mohou infikovat stejného hostitele, na kterém vznikly (sóju), během téže sezóny. Epidemie se může rychle rozvinout z pouhých několika puchýřků, protože puchýřky produkující spory se vytvářejí již za 7 až 10 dní po infekci a každý puchýřek může produkovat stovky urediniospor. Ve starých lézích se vytvářejí teliospory, které však v přírodě zřejmě neklíčí, a není znám žádný náhradní hostitel ani aecia či spermogonia.
Bez klíčení teliospor nemůže dojít k pohlavnímu rozmnožování. Nedostatek pohlavního rozmnožování by měl omezit variabilitu rzi, ale přesto existuje značná variabilita. P. pachyrhizi s ohledem na virulenci. To omezuje používání jednotlivých genů pro rezistenci u sóji, protože v krátké době vznikají nové izoláty patogenu, které gen rezistence překonávají. Není známo, jak tato variabilita vzniká v P. pachyrhizi. Rez pšeničná pruhovaná , Puccinia striiformis, má podobný životní cyklus jako P. pachyrhizi bez funkčního teliálního stadia, a tedy bez pohlavního rozmnožování, ale má mnoho ras. Je možné, že každý gen rezistence je natolik specifický, že jediná mutace ve správném genu houby jí umožňuje být virulentní na hostitelích s novým genem rezistence.
Epidemiologie
Epidemie rzi sójové začínají příchodem inokula (urediniospor) přenášeného vzduchem. Tento patogen je mezi rzí jedinečný, protože má mnoho alternativních hostitelů, kteří mohou sloužit jako zdroje inokula. Alternativní hostitelé jsou jiné rostliny, které se mohou nakazit stejným patogenem, ale nejsou nutné k dokončení životního cyklu patogenu. Alternativní hostitele nelze zaměňovat s alternativním hostitelem, což je jiná rostlina než hlavní hostitel, která je potřebná k dokončení životního cyklu patogenu. V oblastech bez mrazů, jako je Jižní Amerika, Střední Amerika, Karibská pánev, jižní Texas a Florida, může být zdrojem inokula blízká samovolně rostoucí sója, kudzu nebo jiný alternativní hostitel. V oblastech, kde se vyskytují mrazy, jako například na středozápadě Spojených států, musí být inokulum zavlečeno ze zdrojů přezimování, které mohou být vzdáleny stovky kilometrů.
K opětovnému zavlečení obligátních patogenů do vzdálené oblasti dochází u několika dalších chorob, jako je rez stonková pšenice a plísně, např. plíseň modrá na tabáku. Protože spory P. pachyrhizi jsou citlivé na ultrafialové záření, dochází pravděpodobně k dálkovému pohybu těchto spor rzi v bouřkových systémech, kde mraky chrání spory před sluncem. Jakmile životaschopné spory dopadnou na povrch listů vhodného hostitele, infekce a následný rozvoj epidemie závisí na podmínkách prostředí. Obecně dochází k infekci, když jsou listy vlhké a teplota se pohybuje mezi 8 °C a 28 °C, přičemž optimum je 16 °C až 28 °C. Při teplotě 25 °C dochází k určité infekci již po 6 hodinách vlhkých listů, optimální doba je však 12 hodin. Po infekci se mohou během 7 až 8 dnů objevit léze a pustuly s urediniosporami a další infekční cyklus může začít.
Tento krátký životní cyklus znamená, že za vhodných podmínek může epidemie rzi sójové rychle narůst z téměř nezjistitelných hodnot na velmi vysoké. Epidemie rzi sójové může během jednoho měsíce přejít z úrovně pod detekovatelnou úrovní k defoliaci. Může se zdát, že epidemie postupuje ještě rychleji, protože rané infekce se objevují ve spodní části koruny a je obtížné je najít. Epidemie rzi sójové ovlivňuje kromě prostředí také stáří rostlin. Obvykle se na sóji objevují rzi až po odkvětu, pokud se na začátku sezóny nevyskytuje vysoká hladina inokula. Může to být způsobeno větší náchylností rostlin ke rzi, když hostitel vstoupí do reprodukční fáze, může to být proto, že v nižších částech koruny jsou spory více chráněny před UV zářením, nebo to může být proto, že se podmínky v koruně stávají vlhčími, jak se koruny uzavírají. V každém případě se léze mohou tvořit v kterékoli růstové fázi, ale k výraznějšímu nárůstu choroby dochází až po odkvětu.
V programu FieldClimate jsou popsány a vypočteny dva modely pro rez sóji:
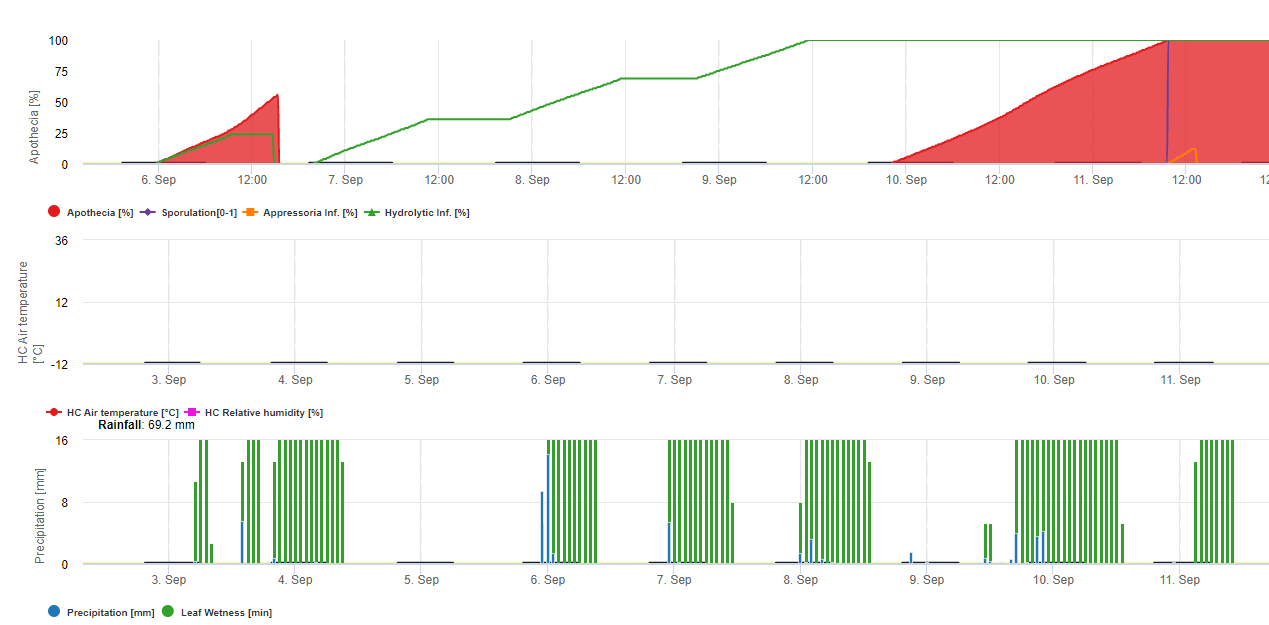
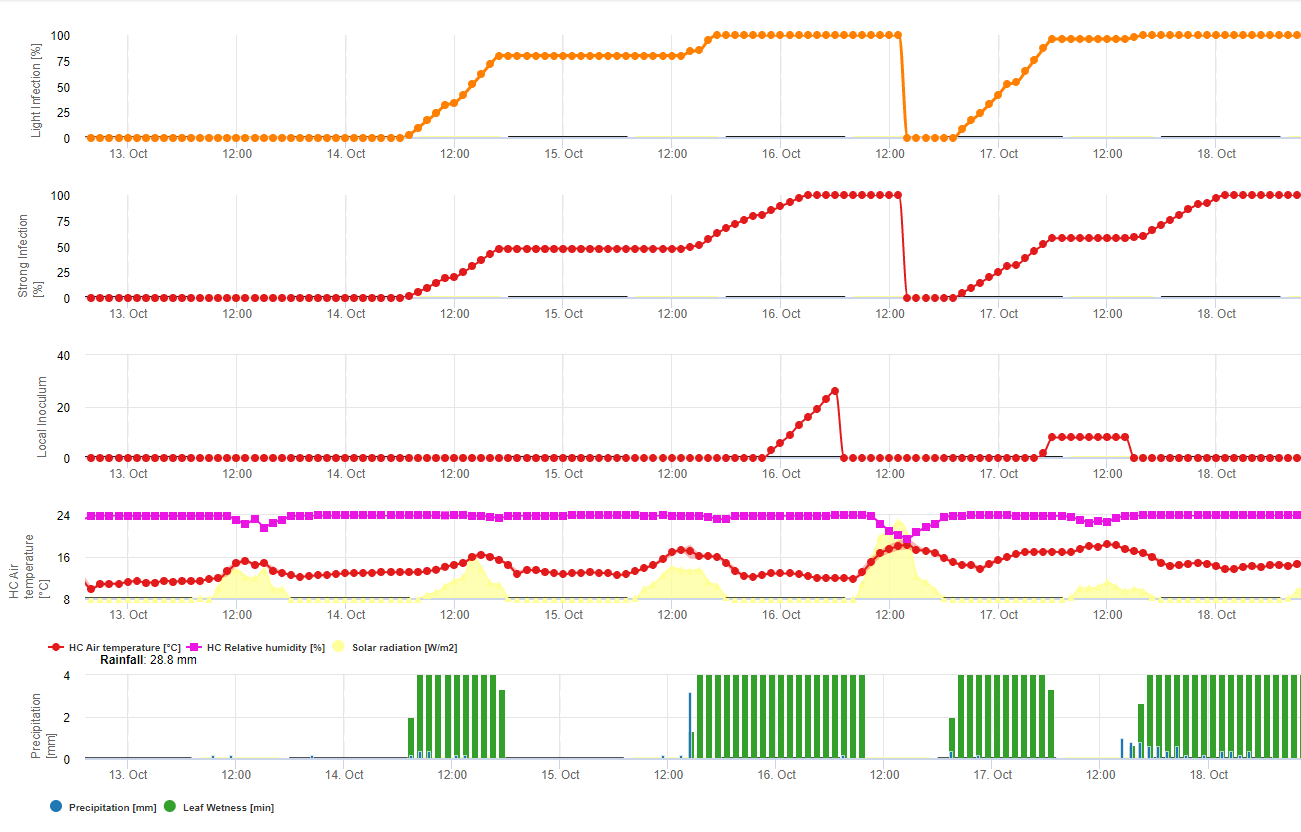
Model 1: Hlavními faktory pro výpočet infekce a závažnosti rzi sójové jsou vlhkost listů a teplota (pozor, infekce musí být 100%, než se začnou počítat stupně závažnosti (od 1-3)) Listy musí být vlhké a teplota musí být mezi 8 a 28 °C - infekce se začne počítat (součet 7200 °minut je 100%). Pokud je infekce 100% a v závislosti na teplotě a délce vlhnutí listů se začnou počítat třídy závažnosti od 1 do 3.
Model 2: Zde je do výpočtu rzi sójové zahrnuto mnohem více faktorů: Teplota, vlhkost listů, relativní vlhkost vzduchu, srážky a sluneční záření (houbové hyfy jsou schopny pronikat do žaludků během dne).
Když infekce rzí sójovou dosáhla 100%, začíná se počítat závažnost. Pokud je obojí (infekce i závažnost 100%), předpokládá se výskyt (usazení?) lokálního inokula.
Literatura:
- Ivancovich, A. y Botta, G. 2002. La roya de la sojo en la Argentinia. EEA Pergamino. RevistadeTEchnologia, Agropecuaria Vol. 7 (21), pp 16-17.
- Melching, J. S., Dowler, W. M., Koogle, D. L. a Royer, M. H. 1989. Vliv délky trvání, frekvence a teploty vlhkých období listů na výskyt rzi sójové. Plant Dis. 73:117-122.
- X.B. Yang, E.M. Del Ponte a A.P. Dias. Poznání rizika výskytu rzi sójové na základě srovnání s Brazílií. Department of Plant Pathology, Iowa State University.
- Yang, X. B. 1995. Hodnocení a řízení rizika výskytu rzi sójové. Sborník ze semináře o rzi sójové, 9.-11. srpna 1995. J. B.Sinclair a G. L. Hartman, eds. National Soybean Research Laboratory, Urbana, IL.
Sklerotiniová hniloba stonků neboli bílá plíseň sóji je onemocnění způsobené houbou Sclerotinia sclerotiorum. Tato choroba může způsobit výrazné snížení výnosu semen nebo dokonce úplně zničit úrodu, pokud jsou sójové boby zasazeny do napadené půdy, kde je hustý porost a dlouhotrvající vlhké počasí. Ke ztrátám výnosů obvykle dochází, pokud je výskyt choroby 15 % nebo vyšší. Ve většině let v celé Severní Dakotě představuje sklerotiniová hniloba stonků pro pěstitele pouze menší až střední problém a choroba se zřídka vyskytuje v letech sucha. Vlhké počasí je hlavním faktorem rozvoje choroby. U zavlažované sóji se jedná o přetrvávající problém. Kromě snížení výnosu osiva má choroba za následek také sníženou kvalitu osiva a osivo kontaminované černými sklerotiemi houby. Kontaminace osiva může být vážným problémem pro vyvážené osivo, protože může vést k odmítnutí partie osiva v zahraničních vstupních přístavech. Kromě toho mohou sklerocia vrácená do půdy ovlivnit další plodiny v osevním postupu. Pěstitelé mohou sklerotiniovou hnilobu stonků zvládnout díky pochopení patogenu a cyklu choroby.
Příznaky
Příznaky se obvykle projeví až v okamžiku, kdy se koruny plodin mezi řádky uzavřou a vytvoří se vlhké mikroklima. Prvními pozorovanými příznaky jsou obvykle vadnutí a usychání listů a následné odumírání rostlin. Podrobná prohlídka pod korunami rostlin odhalí na stoncích, listech nebo luscích bílý myceliální porost (vlákna houby). Léze se vyvíjejí na hlavních stoncích a bočních větvích. Nakonec léze opásají stonky a části rostliny nad nimi odumírají. Stonky vypadají vybělené a někdy roztřepené v důsledku pokročilé hniloby. Z bílého mycelia rostoucího na rostlinném pletivu se vytvoří velká černá sklerocia různých tvarů a velikostí. Sklerocia se tvoří také v jádru stonků a mají charakteristický válcovitý tvar. Semena v nemocných luscích jsou obvykle scvrklá a mohou být napadena houbou nebo nahrazena černými sklerotiemi. Při sklizni infikovaných rostlin je osivo obvykle kontaminováno sklerotiemi.
Biologie patogenů
Tato houba má rozsáhlý hostitelský areál čítající více než 370 druhů rostlin a způsobuje choroby na široké škále plodin, jako je slunečnice, suché fazole, řepka, brambory, vojtěška, pohanka, lupina, hořčice, topinambur, světlice barvířská, čočka, len, hrách a mnoho druhů zeleniny. V Severní Dakotě je vzácné, že tento patogen způsobuje vážné škody na některých z těchto plodin, jako je len a brambory. Existuje také mnoho běžných hostitelů z řad širokolistých plevelů, jako jsou blatouch bahenní, jitrocel kopinatý, prasetník, bodlák kanadský a hořčice divoká. Houba, která způsobuje bílou plíseň na sóji, je stejná jako ta, která způsobuje bílou plíseň neboli sklerotiniózu slunečnice, suchých fazolí, řepky a dalších plodin. Sclerotinia sclerotiorum přezimuje hlavně jako sklerocia v půdě. Sklerocia vyklíčí a vytvoří malé hnědé až hnědé houby zvané apothecia (asi osmina až čtvrtina palce v průměru). Ty produkují spory označované jako askospory, které iniciují onemocnění sóji a dalších náchylných plodin.
Cyklus onemocnění
Vlhkost a kvetení jsou rozhodujícími faktory pro rozvoj choroby. Choroba se obvykle nevyskytuje před uzavřením koruny, protože hustá koruna podporuje nízké teploty a vlhké mikroklima kolem stonků a udržuje vysokou vlhkost půdy po dešti nebo zavlažování. Iniciace choroby je rovněž úzce spojena s kvetením. Po sedmi až čtrnácti dnech vysoké půdní vlhkosti sklerocia v horních několika centimetrech půdy vyklíčí a vytvoří houbovitá apothecia. Z jednoho sklerocia může vzniknout několik apothecií. Apothecia násilně vyvrhují své askospory do vzduchu, odkud jsou vzdušnými proudy přenášeny na rostliny sóji. Nejdůležitějším zdrojem askospor jsou apothecia produkovaná na poli, ale askospory mohou být zaváté ze sousedních nebo blízkých polí. Jedno apothecium může během několika dní vyprodukovat obrovské množství askospor. Askospory přežívají krátkou dobu na rostlinných pletivech, ale nepřezimují. Askospory potřebují ke klíčení a růstu vrstvu vody a potravní základnu, jako jsou odumřelá nebo odumírající květní pletiva, než infikují rostlinu. Květní pletiva jsou nejdůležitějším základem pro iniciaci infekce. Infekce často začínají v paždí stonků, kde opadávají a ulpívají odumírající květní pletiva. K infekci může dojít také přes rány způsobené krupobitím nebo jiným poraněním. Film vody na povrchu rostliny podporuje rozvoj lézí a zvyšuje míru poškození tkání. Počáteční vývoj choroby obecně vyžaduje více než 40 hodin nepřetržitého vlhka na povrchu rostliny, ale jakmile se choroba rozvine, kratší období vlhka umožní rozvoj lézí. To je důvod, proč je choroba spojena s dlouhými obdobími zamračeného, vlhkého a deštivého počasí. Jakmile povrch rostlin vyschne, postup choroby se zpomalí. Chladné teploty mezi 59 a 75 stupni Fahrenheita podporují rozvoj choroby. Čím větší je hustota rostlinného porostu, tím příznivější jsou podmínky prostředí pro chorobu. Sója s pevným výsevem a polehlé porosty jsou proto náchylnější k rozvoji chorob. Sklerocia se vytvoří při růstu mycelia v rostlinných pletivech a na nich. Tato sklerocia během sezóny nevyklíčí a nevytvoří další apotécia, ale vrátí se do půdy při sklizni a zpracování půdy a přezimují, aby se stala inokulem (zdrojem infekční houby) pro budoucí náchylnou plodinu. Sklerocia jsou velmi odolné struktury a přežívají v půdě po dlouhou dobu.
Zdroj: http://www.ndsu.edu/pubweb/~bernelso/soydiseases/sclerotinia.shtml
V programu FieldClimate se model pro sklerotinii počítá v závislosti na období dešťů, relativní vlhkosti a teplotě a také na vlhkosti listů. Během dlouhých vlhkých období se doporučuje infekce prostřednictvím založení apresoria houbovým patogenem. Další způsob infekce se nazývá "hydrolytická infekce". Tento způsob je založen na uvolňování hydrolytických enzymů, které postupně degradují rostlinnou kutikulu, střední lamely a primární a sekundární buněčné stěny a celou rostlinu. Podrobně popsaný model na této stránce.