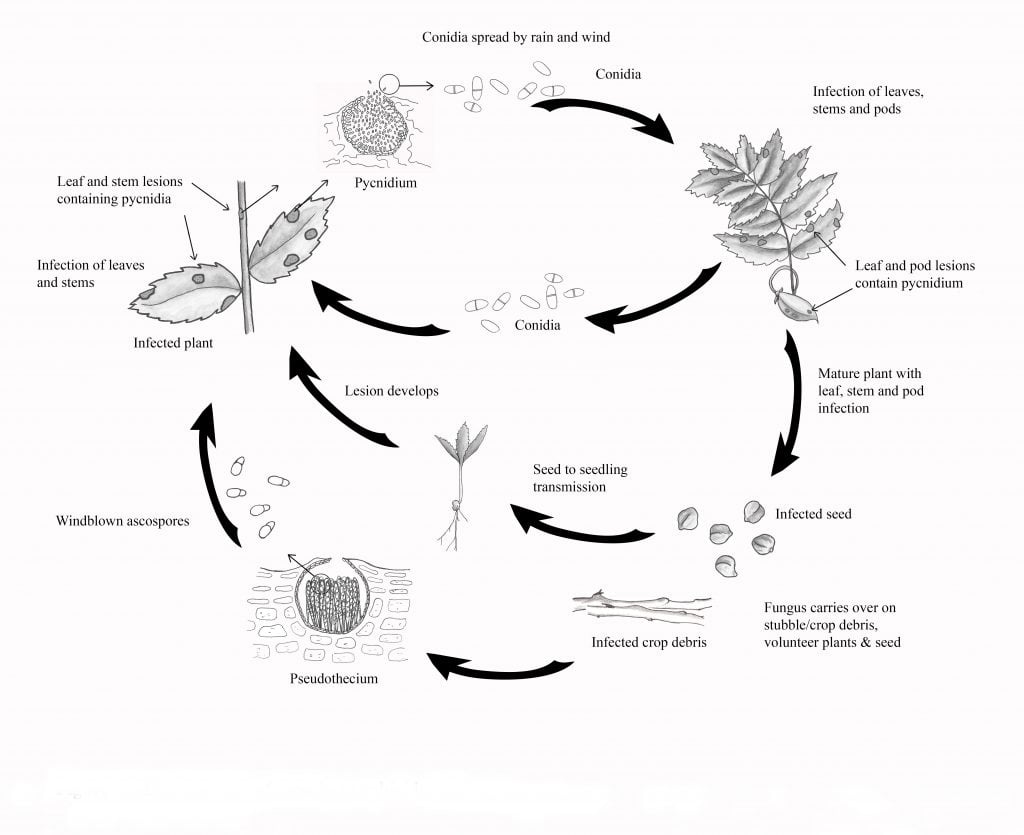
Životní cyklus
Patogen přezimuje na infikovaných rostlinných zbytcích a osivu. Infikované osivo může hrát důležitou roli jak při zavlečení Ascochyta Blight do nových oblastí, tak i při časném rozvoji choroby, protože patogen se snadno přenáší ze semen na sazenice. Na rostlinných zbytcích se mohou vytvářet jak nepohlavní spory (konidie, šířící se deštěm), tak pohlavní spory (askospory, šířící se větrem). Při pohlavním rozmnožování se koncem podzimu a začátkem jara vytvářejí pseudothecia, v nichž se nacházejí askospory. Vývoj pseudotécií trvá pět až sedm týdnů při dostatečné vlhkosti a mírných teplotách (kolem 10 °C).
Na jaře a začátkem léta uvolňují zralá pseudothecia do vzduchu askospory, které mohou cestovat několik kilometrů a infikovat cílové plodiny. Předpokládá se, že prvotním zdrojem infekce na jaře jsou vzdušné askospory, i když se na ní mohou podílet i konidie rozptýlené deštěm. Po infekci sporami se příznaky začnou projevovat během čtyř až šesti dnů. Časné léze jsou hnědé až tmavě hnědé s tmavě hnědým okrajem. Tři až šest dní po vzniku lézí se objevují tmavě hnědé pyknidy.
Pyknidy jsou často uspořádány do soustředných kruhů a neodírají se o zbytky tkáně. Konidie vytékají z pyknid v lepkavé hmotě výtrusů a šíří se dešťovou sprškou na zdravé části rostlin, kde způsobují nové infekce.
Většina poškození během vegetačního období je důsledkem rychlého vývoje pyknid a konidií za vlhkých podmínek. I malé dešťové přeháňky stačí k rozšíření konidií do nových rostlinných pletiv. Proto se tato choroba nazývá polycyklická, což znamená, že při dostatečné vlhkosti a teplotách (20 až 25 °C) může dojít k více infekčním cyklům během vegetačního období.
Model onemocnění
Modelujeme vývoj pyknid a konidií během růstového období.
Vývoj choroby Ascochyta blight je optimální při teplotách 20 až 21 °C (teplotní rozmezí je 4 °C až 34 °C) a vlhkých podmínkách (vysoká relativní vlhkost a vlhkost listů).
Do modelu jsou implementovány dva kroky cyklu onemocnění: 1. Sporulace (vývoj pyknid, uvolňování konidií), tedy produkce nových infekčních jednotek, a 2. optimální podmínky pro další infekci těchto konidií. Pro infekci nového rostlinného pletiva byly tedy nejprve dány optimální podmínky pro sporulaci (100%) a poté infekční křivka dosáhla 100%.
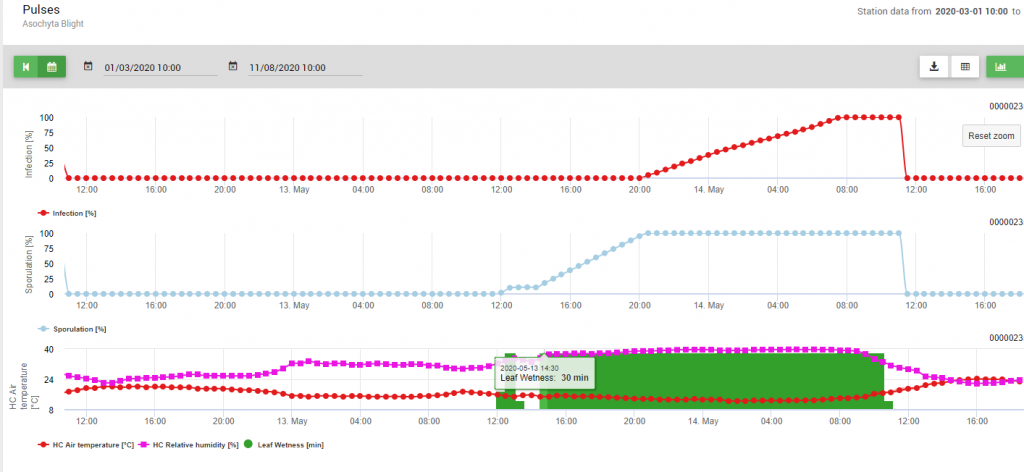
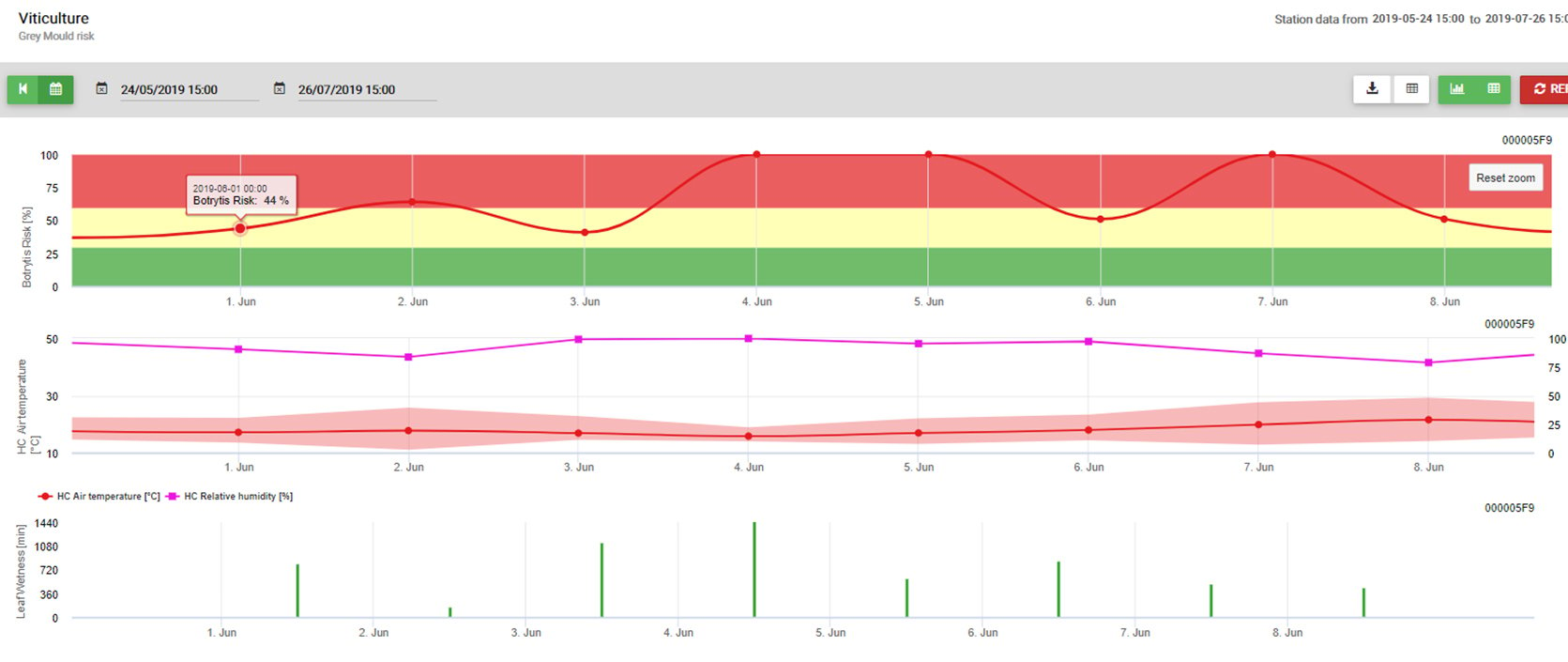
Vzhledem k optimálním podmínkám (vlhkost listů, vysoká relativní vlhkost a teplota kolem 15 °C po dlouhou dobu) byla sporulace pyknid stanovena 13. května ve 20:00 (modrá linie, 100%). Podmínky byly stále optimální pro zahájení postupu infekce (červená čára) a optimální podmínky pro infekci byly zjištěny 14. května v 8:00 ráno (dosaženo 100%). V závislosti na strategii ochrany rostlin tedy měla být zohledněna profylaktická měření již před výskytem infekce nebo při použití kurativních měření krátce po zjištění infekce (100%).
Literatura:
Botrytis cinerea je nekrotrofní houba, která napadá mnoho druhů rostlin, ačkoli jejími nejvýznamnějšími hostiteli jsou vinné hrozny.
Ve vinařství je běžně známá jako botrytická hniloba hroznů, v zahradnictví se obvykle nazývá plíseň šedá nebo plíseň šedá.
Tato houba způsobuje na hroznech dva různé druhy infekcí. První z nich, šedá hniloba, je důsledkem trvale vlhkých nebo mokrých podmínek a obvykle vede ke ztrátě napadených hroznů. Druhá, ušlechtilá hniloba, vzniká, když po vlhčích podmínkách následují sušší, a může mít za následek vznik výrazných sladkých dezertních vín, jako je Sauternes nebo tokajské Aszú. Název druhu Botrytis cinerea je odvozeno z latinského "hrozny jako popel"; ačkoli je to poetické, "hrozny" odkazují na shluk houbových výtrusů na jejich konidioforech a "popel" pouze na šedavou barvu výtrusů. Houba se obvykle označuje jménem anamorfa (nepohlavní forma), protože pohlavní fáze je pozorována jen zřídka. Teleomorfa (pohlavní forma) je askomycet, Botryotinia cinerea.
Biologie B. cinerea
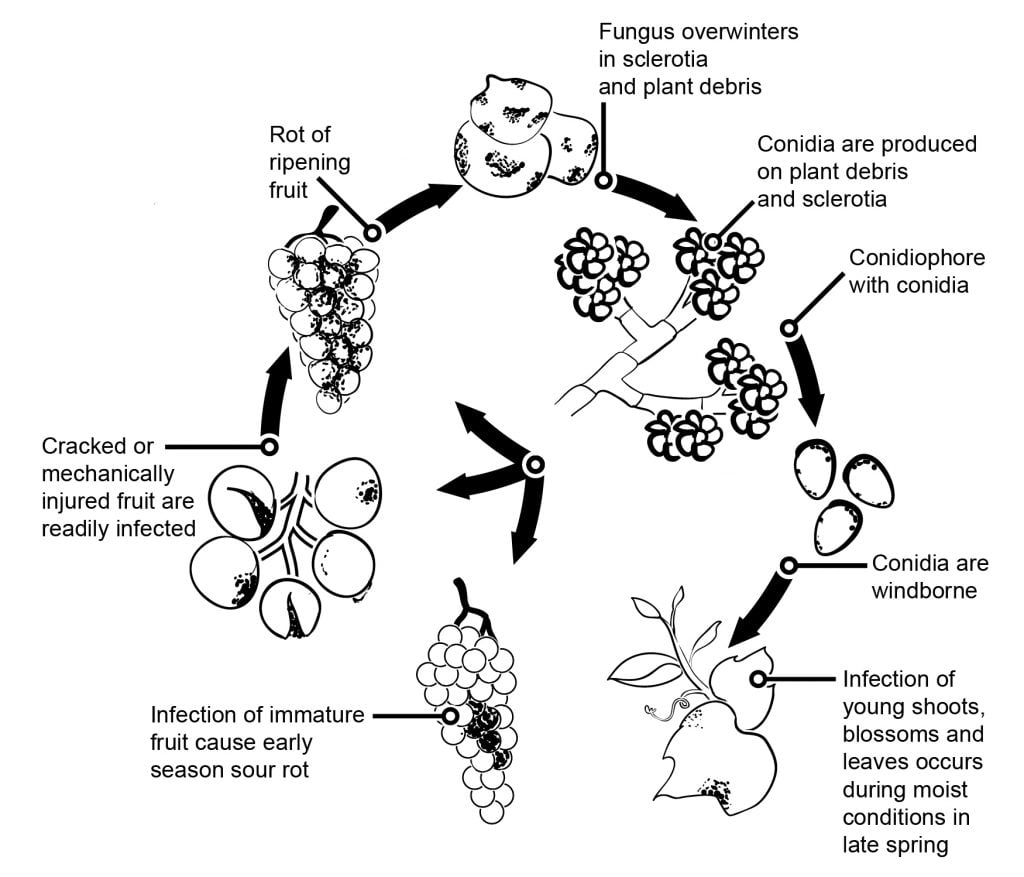
V případě ovoce a hroznů mají na epidemiologii plísně šedé velký vliv květové infekce, po nichž následuje latence. Lze oddělit několik cest infekce z květů do plodů. U hroznů, kiwi a malin byla postulována infekce přes tyčinky do vaječníku. Ve vaječníku zůstává patogen latentní, což je zřejmě důsledkem předem vytvořené obranné strategie hostitele (podobně jako obsah resveratrolu v mladých bobulích hroznů). U vinné révy, kiwi a jahodníku byla zjištěna významná infekce přes tyčinku, okvětní lístky nebo kališní lístky. Studie ukázaly, že u vinných hroznů B. cinerea může infikovat tyčinky a prorůstá bazipetálně, aby infikoval receptakulum, a poté prorůstá systémově do stopky a cévních tkání v bobulích.

Šestiletý výzkumný projekt v Kapské oblasti ukázal, že bobule hroznů mohou být infikovány přes stopku a čočky na stopce a v dolní části stopky. Infekce stopky je možná i v období květu. Později tato tkáň zvyšuje odolnost proti B. cinerea infekce.
Další způsoby infekce předpokládají saprofytický růst patogenu na zbytcích květů a pozdější infekci bobulí, když se náchylnost zvyšuje s dozráváním nebo poškozením bobulí hmyzem či krupobitím. Další tezí je předpoklad hromadění konidií v plodech během léta a infekce náchylných bobulí později v sezóně. Infekce dozrávajících plodů konidiemi je možná z jakéhokoli zdroje inokula. S největší pravděpodobností vzniká nízký počet latentně infikovaných bobulí, které vykazují rozsáhlou sporulaci, když se náchylnost bobulí zvyšuje s jejich zralostí. Lze předpokládat, že bobule se stanou náchylnými od cukernatosti 6%.
V případě kiwi mají velký vliv podmínky sběru na výskyt B. cinerea. Ovoce sklízené na mokrém povrchu se může nakazit B. cinerea na mikrozranění, která vznikla v prstech sběračů.
Při praktické kontrole B. cinerea musíme oddělit dvě důležitá infekční období: Květ a stárnutí. 1) Během kvetení máme náchylné mladé plody, kde po infekci následuje období latence. 2) Zatímco infekce na zralých (senescentních) plodech povede k symptomům bez období latence. Význam infekce během kvetení se u hroznů může v jednotlivých sezónách a regionech měnit. U ovoce, u kterého musíme počítat s určitou trvanlivostí (stolní hrozny, kiwi nebo jahody), se příznaky projevují při skladování v chlazených podmínkách v obchodech nebo skladech. Chemická ochrana révy vinné vykazující dobrou odolnost proti B. cinerea během kvetení nebude mít žádné hospodářské výsledky. Proto je třeba při rozhodování o žádosti o ošetření zohlednit všechny podmínky rizika a pravděpodobnosti infekce, citlivost plodů a dobu skladovatelnosti, podmínky skladování proti chorobám, které se vyskytují v plodinách. Botrytis cinerea během kvetení.
U peckovin infekce B. cinerea se vyskytuje hlavně během kvetení. V této době se provádí ošetření proti Monilina spp. jsou brány v úvahu, které také infikují Botrytis cinerea infekce.
Model B. cinerea a praktické využití
Potřebné senzory: Vlhkost listů, teplota, relativní vlhkost vzduchu
Botrytis cinerea je fakultativní parazit. Roste i na odumřelém rostlinném materiálu. Z tohoto důvodu se vždy vyskytuje ve vinicích a sadech. Botrytis cinerea je vázána na vlhké klima. K infekci potřebuje velmi vysokou relativní vlhkost vzduchu nebo přítomnost volné vody (senzor: vlhkost listů, relativní vlhkost vzduchu). Houba není schopna infikovat zdravý dospělý rostlinný materiál sporami. K infekci dochází na mladých výhonech révy během delších vlhkých období nebo na výhonech poškozených krupobitím.
Model používá k výpočtu rizika infekce následující korelaci mezi délkou trvání vlhkosti listů a teplotou.
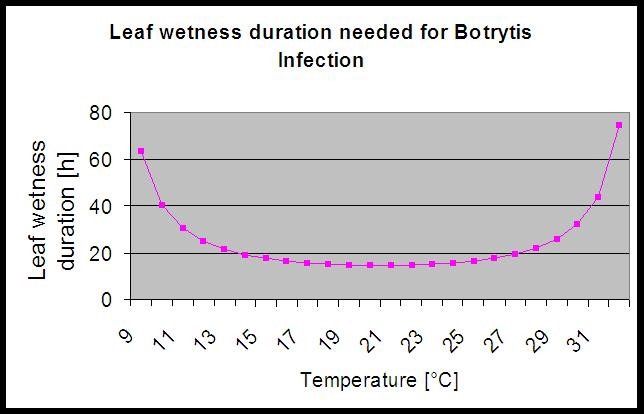
Na stránkách Botrytis Výsledkem modelu rizika je hodnota rizika 0 až 100%. Tato hodnota udává tlak B. cinerea v té době. Pokud máme hodnotu 100%, znamená to, že několikrát proběhlo dostatečně dlouhé období vlhkosti, aby došlo k infikování náchylného pletiva (počítáme tzv. "mokré body" (pole mezi vlhkostí listů, teplotou s maximem původně 38400 bodů (začátek sezóny, který vykazuje riziko 30%). Po tomto období každé vlhké období s přibližně 4000 vlhkými body (pole) zvyšuje riziko o 10% nebo naopak každé suché období snižuje riziko o 1/5 původní hodnoty. Aplikace proti
Žádost proti B. cinerea závisí na druhu ovoce a cíli produkce.