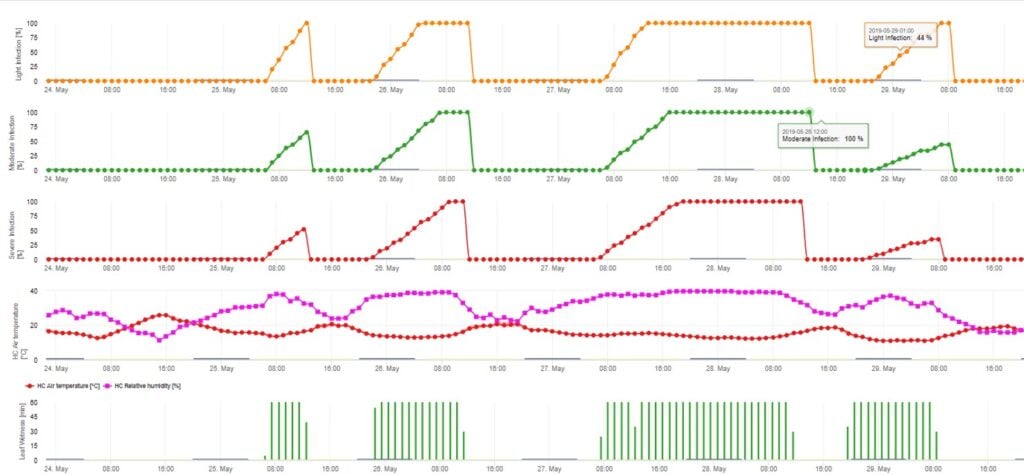
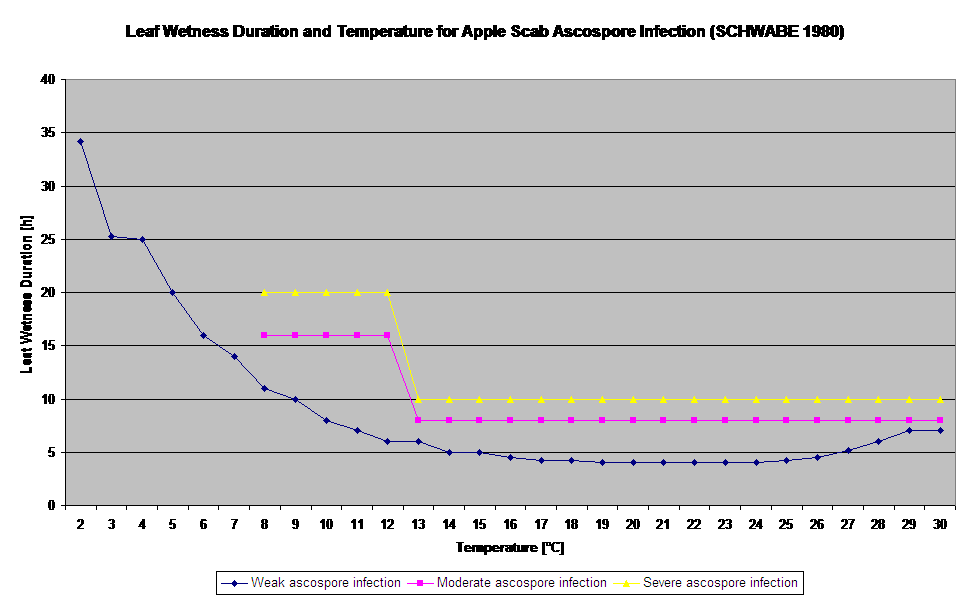
Klíčení a penetrace askospor Venturia inaequalis na listech nebo plodech závisí na vlhkosti listů (vlhké listy nebo relativní vlhkost >80% (udržuje úroveň infekce až do 80% r.h.) a teplotě vzduchu (nad 2 °C). Vyšší teploty vedou za vlhka k rychlejšímu pronikání, zatímco při nižších teplotách je zapotřebí delší období vlhka listů. Tento vztah poprvé publikovali MILLS a LAPLANTE (1945). Náš výpočet je proveden na základě publikací SCHWABEHO (1980). SCHWABE popsal závažnost infekce strupovitosti v závislosti na teplotě. Tato pozorování jsou ve shodě s našimi vlastními pozorováními.
Model infekce askosporami vypočítává průběh infekce v závislosti na vlhkosti listů a teplotě pro slabé, středně silné a silné infekce. Rozdělení předpovědi do tří "tříd závažnosti" je pro modely strupovitosti jabloní velmi tradiční. Průběh infekce se zobrazuje graficky pro hodinové hodnoty. K infekci již došlo, jakmile křivka průběhu slabé infekce dosáhne hodnoty 100%. Středně silná nebo silná infekce je dokončena, pokud křivky průběhu pro tyto třídy závažnosti dosáhnou (100%). V praxi nebo ve skutečnosti často infekce dosáhne 70% nebo 80% brzy ráno a v závislosti na aktuálním počasí budete moci poznat, zda tato infekce skončí, nebo ne. Zobrazení denních maximálních hodnot křivky průběhu infekce poskytuje přehled o očekávaných termínech infekce v sezóně.
Primární použití modelu
Pro trh je naprosto důležité mít plody jablek prosté chorob, aby bylo možné získat dobrou cenu. Proto se pěstitelé snaží ve svých sadech vůbec netolerovat strupovitost. Model by měl být implementován do programu léčebné aplikace (aplikace postřiků obsahujících sloučeniny cyprodinilu nebo pyrimethanilu v době před květem), když model poukázal na infekci (bez ohledu na to, zda slabou nebo silnou). V období květu a na počátku období po odkvětu se nejčastěji vyskytují silné infekce a všechna vlhká období byla pokryta ochrannými postřiky. Dále rostlinná pletiva velmi rychle rostou, proto může ochrana působit pouze 4 dny. V takovém případě může být nutný i léčebný postřik po neúplném infekčním období. V tomto případě je však důležité, zda se jednalo o slabou nebo silnou infekci a také zda byl nízký nebo vysoký počet askospor připravených k infekci.
Literatura
- Schwabe WFS, 1980. Požadavky na vlhkost a teplotu pro infekci listů jabloní Venturia inaequalis v Jižní Africe. Phytophylactica, 69-0.
- Schwabe WFS, 1980. Počasí příznivé pro infekci strupovitostí jabloní v Jižní Africe. Phytophylactica 12, 213-217.
- Mac Hardy W.E., 1996. Apple scab biology, epidemiology and management. St. Paul, MN: Am. Phytopathol. Soc.: 545.
Sekundární infekce konidiemi
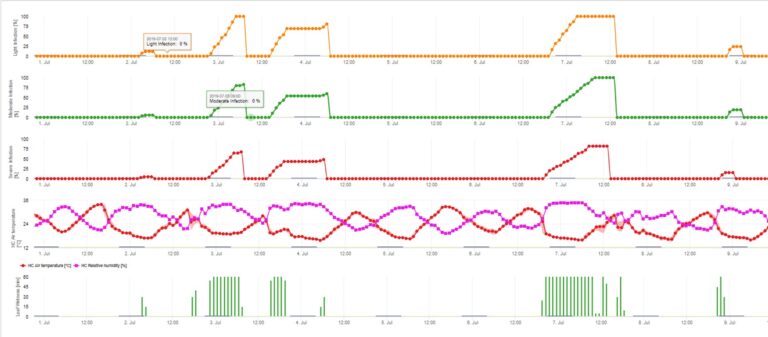
Infekce strupovitosti jabloní konidiemi (asexuální) závisí na stejných faktorech jako infekce způsobené askosporami. Rozdíl spočívá v tom, že konidie se mohou uvolňovat za světla i za tmy a k infekcím konidiemi dochází převážně v letním období.
V závislosti na teplotě vzduchu potřebují konidie Venturia inaequalis delší nebo kratší období vlhkosti listů, aby vyklíčily a pronikly do listů nebo plodů jabloně. Tento vztah poprvé publikovali MILLS a LAPLANTE (1945). Naše výpočty jsou provedeny na základě publikací SCHWABEHO (1980). Zatímco Mills a LAPLANTE (1945) pouze předpokládali, že konidie potřebují určitou dobu vlhka v listech, SCHWABE (1980) zahrnul do procesu infekce také význam teploty.
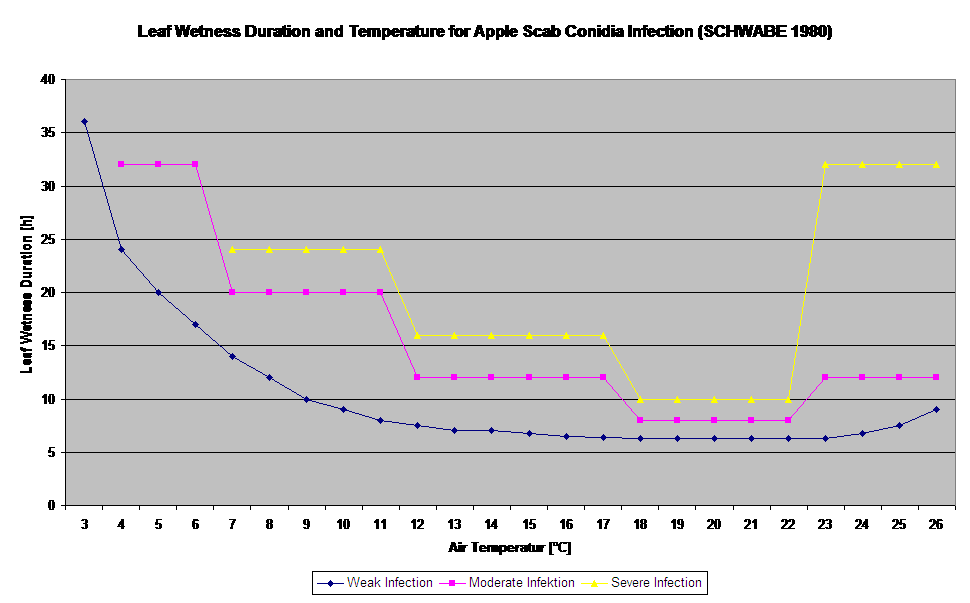
Model infekce konidiemi počítá průběh infekce v závislosti na vlhkosti listů a teplotě pro slabé, středně silné a silné infekce. Rozdělení předpovědi do "tří tříd závažnosti" je pro modely strupovitosti jabloní velmi tradiční. Průběh infekce se zobrazuje graficky pro hodinové hodnoty. K infekci již došlo, jakmile křivka průběhu slabé infekce dosáhne hodnoty 100%. Středně silná nebo silná infekce je dokončena, pokud křivky průběhu pro tyto třídy závažnosti dosahují 100%. V praxi nebo ve skutečnosti často infekce dosáhne 70% nebo 80% brzy ráno a v kombinaci s aktuálním počasím budete schopni poznat, zda tato infekce skončí nebo ne. Zobrazení denních maximálních hodnot křivky průběhu infekce poskytuje přehled o očekávaných termínech infekce v sezóně.
Literatura:
- infekce Venturia inaequalis s uvedením dat infekce askosporami a konidiemi (SCHWABE, W. 1980)
- Mac Hardy W.E., 1996. Apple scab biology, epidemiology and management. St. Paul, MN: Am. Phytopathol. Soc.: 545.
Grafické znázornění modelu
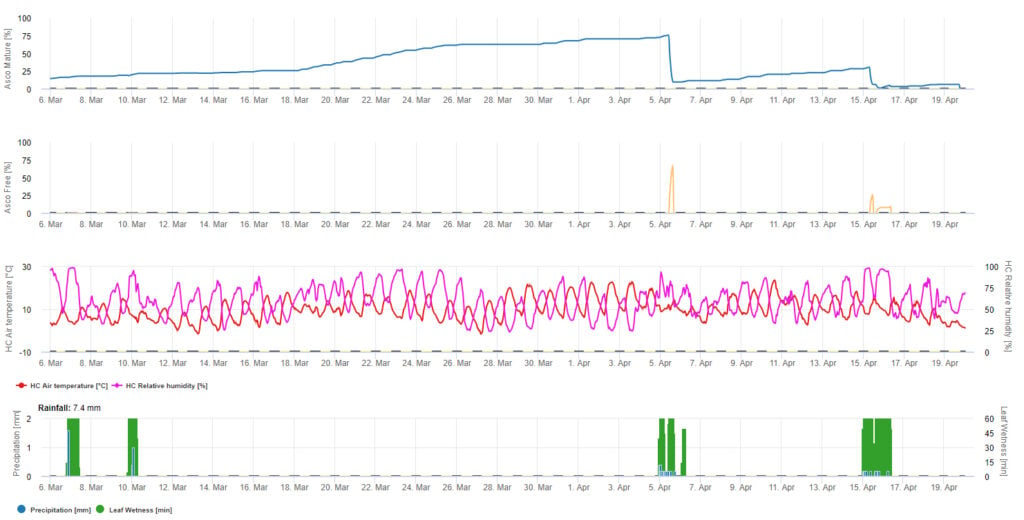
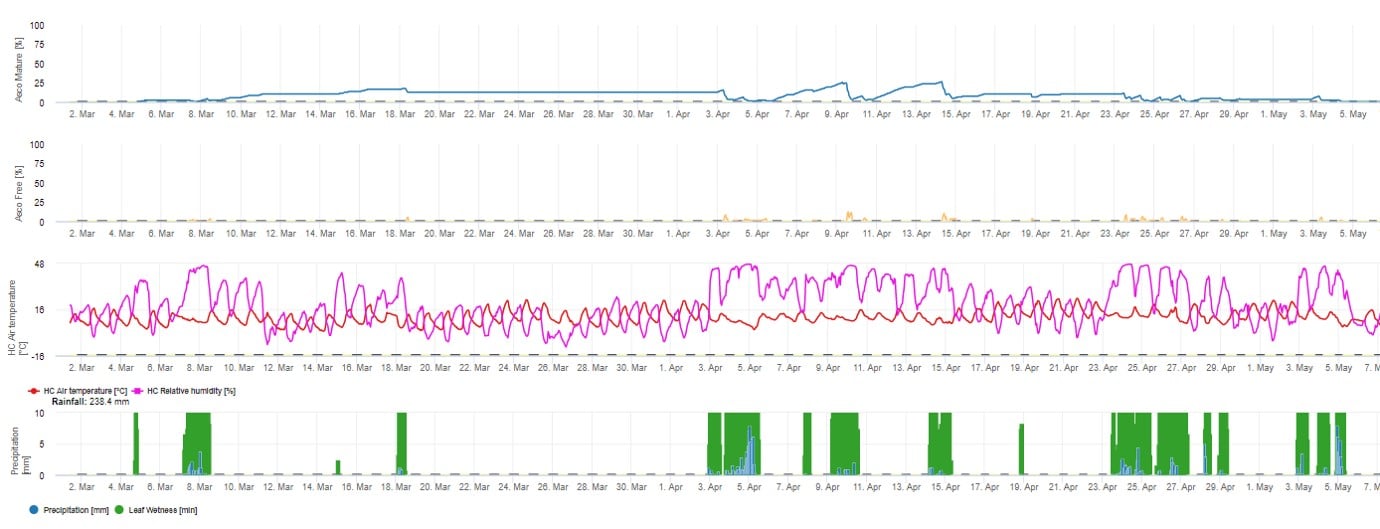
Pro grafické znázornění zrání, tvorby a vypouštění askospor jsou použity denní hodnoty modelu. Z něj vyplývá, že období vlhka listů vedou k uvolňování askospor a ke křivkám zralých a uvolněných askospor. Závažnost uvolňování a počty zralých askospor nelze hodnotit v absolutních hodnotách. Hodnoty mezi 0 a 100% udávají relativní množství zralých a uvolněných askospor v závislosti na klimatických podmínkách.
1. Zrání, tvorba a vypouštění askospor:
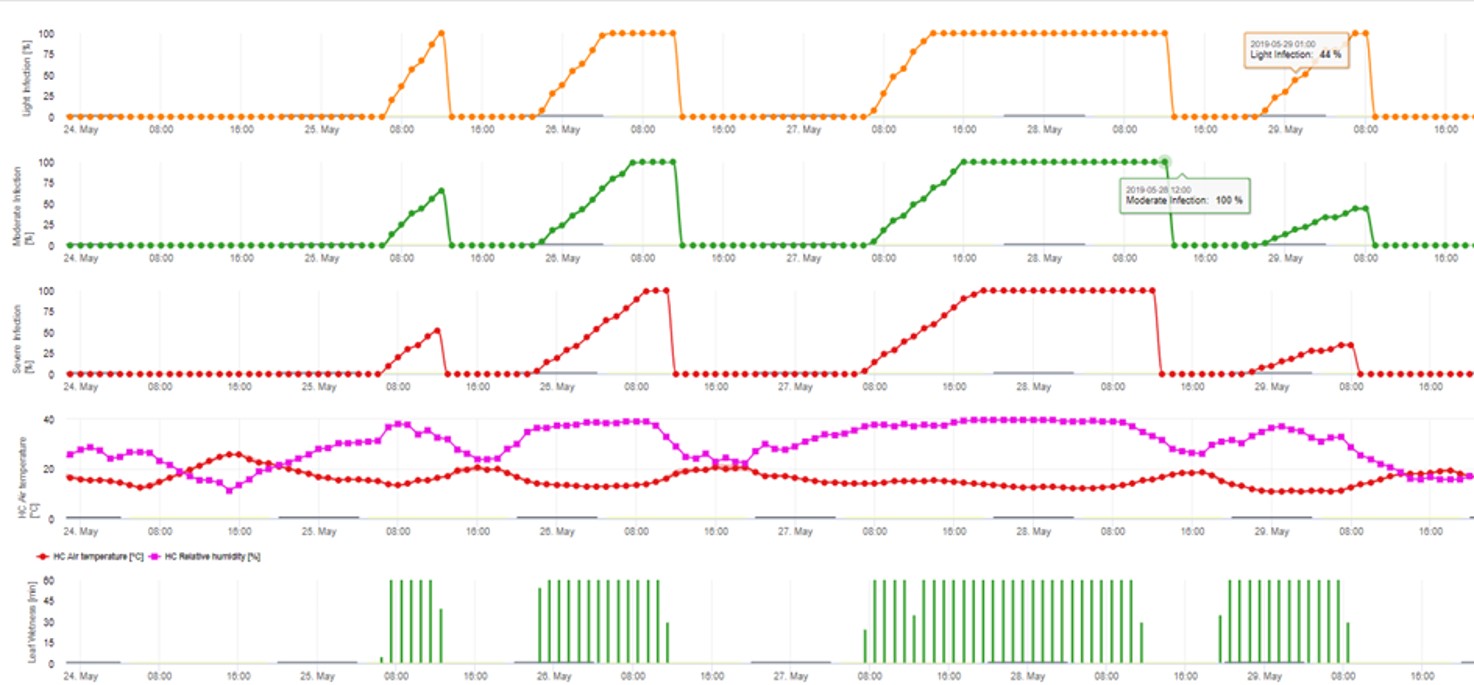
2. Infekce askospor zobrazená ve třech různých třídách závažnosti (slabá: oranžová, středně silná: zelená, silná: červená). Kdykoli bylo dosaženo infekce 100%, byly stanoveny optimální podmínky pro infikování rostlinného pletiva askosporami. Je tedy třeba vzít v úvahu měření ochrany rostlin (před infekcí profylaktické, po infekci 100% kurativní):
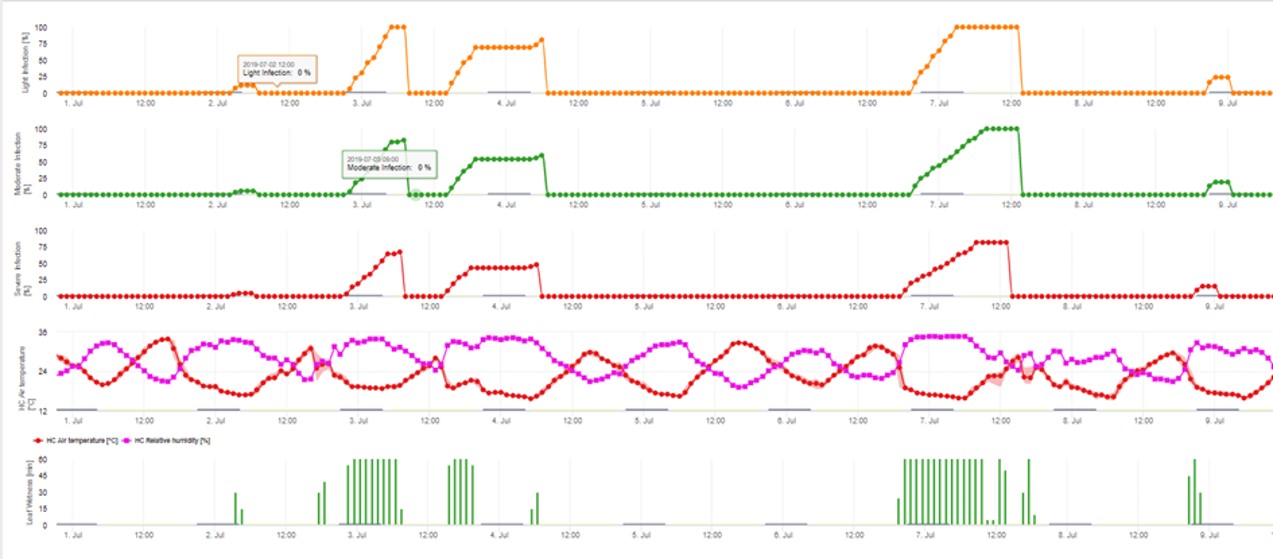
3. Infekce konidiemi se zobrazuje ve třech různých třídách závažnosti (slabá: oranžová, středně silná: zelená, silná: červená). Kdykoli bylo dosaženo infekce 100%, byly stanoveny optimální podmínky pro infikování rostlinného pletiva askosporami. Je tedy třeba vzít v úvahu měření ochrany rostlin (před infekcí profylaktické, po infekci 100% kurativní):
Komerčně pěstovaná jablka musí být bez strupovitosti. Jablka se strupovitostí se prodávají pouze ke zpracování. Proto je cílem všech činností na ochranu rostlin v konvenčním i ekologickém pěstování mít plody bez strupovitosti. Modely, které ukazují vypouštění askospor z jabloně a infekci askosporami/konidiemi, jsou velmi důležitými nástroji k dosažení tohoto cíle.
V konvenčních systémech pěstování se používají dva základní typy fungicidů proti strupovitosti jabloní: a) preventivní přípravky jako Captan, Mancozeb, Dithianone a Strobiluriny nebo b) kurativní přípravky jako Cyprodinil (Chorus) nebo Pyrimethanil (Scala) nebo pro aplikaci v pozdějších teplejších obdobích sezóny DMI fungicidy. Ve skutečnosti většina pěstitelů uplatňuje preventivní strategii. Nicméně praktická preventivní strategie není schopna jabloně zcela ochránit, protože jabloně rostou a vyvíjejí květy, plody a listy. Proto preventivní postřik chrání pouze po dobu 4 až 7 dnů v závislosti na aktuálním růstu stromu. Takto úzké intervaly postřiků nelze zvládnout, proto pěstitelé do managementu postřiků zahrnou své zkušenosti s místním klimatem, předpovědí počasí a modely strupovitosti jabloní. Preventivní postřiky naplánují na základě svých zkušeností a předpovědi počasí. Modely infekce strupovitostí jabloní jim ukáží přesný termín infekce (slabá, střední a silná), stejně jako model výskytu askospor/konidií, a díky svým zkušenostem jsou schopni odhadnout význam infekce. To dává možnost zasáhnout kurativním přípravkem, pokud infekce strupovitostí jabloní proběhla příliš dlouho po posledním preventivním postřiku.
V ekologické produkci jablek se jako nejúčinnější prostředek proti strupovitosti ukázala vápenná síra. Optimální kontroly lze dosáhnout, pokud se postřik provede krátce před infekcí nebo do začátku infekce. Tento postřik je třeba naplánovat na základě předpovědi počasí. Někdy toto optimální období promeškáme a musíme provést postřik do vlhkých listů téměř dokončené infekce strupovitosti. Tím se ještě dosáhne dobré účinnosti. Modely strupovitosti jabloní pomáhají rozhodnout, zda je nutný emergentní postřik do téměř úplné infekce.
Příznaky ohnivé nákazy
Přezimující hniloby, v nichž se vyskytuje patogen ohnivé hniloby, jsou často zřetelně viditelné na kmenech a velkých větvích jako mírně až hluboce propadlé plochy se změněnou barvou kůry, které jsou někdy na okrajích popraskané. Největší počet rakovinných buněk je však mnohem menší a není tak snadné je rozlišit. Vyskytují se na malých větvích, kde v předchozím roce došlo k napadení květů nebo výhonů, a často kolem řezů provedených při odstraňování napadených větví. Vzhledem k tomu, že mnoho těchto rakovin vzniká později v průběhu sezóny, nebývají často silně vpadlé a jen zřídkakdy se na jejich okrajích objevují praskliny kůry. Často jsou také poměrně malé, sahají méně než 2,5 cm, s načervenalou až fialovou kůrou, která může být pokryta drobnými černými plodnicemi hub (nejčastěji Botryosphaeria obtusa, původce černé hniloby jablek).
Příznaky plísně květů se nejčastěji objevují během jednoho až dvou týdnů po odkvětu a obvykle se týkají celého květního úboru, který vadne a odumírá, přičemž u jabloní hnědne a u hrušní zcela černá. Pokud je počasí příznivé pro rozvoj patogenu, lze na květech pozorovat kuličky bakteriálního mazu. Odumírá také odnož nesoucí květní úbor a infekce se může rozšířit do části podpůrné větve a zahubit ji. Špičky mladých infikovaných výhonů vadnou a vytvářejí velmi typický příznak "pastýřského chomáče". U starších výhonů, které se nakazí po vytvoření asi 20 listů, se tento příznak kroucení na špičce nemusí projevit. Jak se infekce šíří po ose výhonu, listy nejprve vykazují tmavé pruhy ve středních žilkách, poté vadnou a hnědnou a zůstávají pevně přichyceny k výhonu po celou sezónu. Stejně jako v případě infekce květů patogen často napadá a ničí část končetiny, která podpírá infikovaný výhon. Prvním příznakem na vodních výhonech a výhonech, které jsou napadeny systémově z blízkých aktivních rakovin, je vznik žlutého až oranžového zbarvení špičky výhonu předtím, než dojde k vadnutí. Kromě toho řapíky a střední žilky bazálních listů na těchto výhonech obvykle odumírají dříve než listy na špičce výhonu.
V závislosti na kultivaru a jeho vývojovém stádiu v době infekce může infekce jediného květu nebo výhonu vést k odumření celé větve, a pokud je napadena centrální lípa nebo kmen stromu, může být během jediné sezóny zničena větší část stromu. Obecně platí, že infekce jakéhokoli typu, které se objeví mezi opadem okvětních lístků a nasazením terminálního pupenu, obvykle vedou k největší ztrátě končetin a stromů. Kromě toho silně strukturované stromy obvykle trpí méně závažnou ztrátou končetin než stromy vyškolené na slabší systémy pro vysokou produktivitu. Pokud dojde k napadení vysoce náchylných podnoží jabloní (M.26, M.9), zůstává velká část kmene a hlavních větví nad spojením roubů velmi typicky bez příznaků, zatímco kolem podnože vzniká výrazná tmavě hnědá rakovina. Jakmile tato rakovina podnože opáše strom, horní část stromu vykazuje do poloviny až konce sezóny příznaky celkového úpadku (špatné zbarvení listů, slabý růst). V některých případech se listy stromů postižených podnožovou hnilobou koncem srpna až začátkem září zbarví do červena, což není nepodobné barvě, která je často spojována s chorobou límcovité hniloby způsobenou houbou přenášenou půdou. U některých stromů s infekcí podnoží se mohou příznaky úpadku projevit až na jaře následujícího roku, kdy lze pozorovat rakovinu, která se rozšiřuje směrem vzhůru do spodní části kmene.
Cyklus onemocnění
Bakteriální patogen způsobující ohnivou skvrnitost přezimuje téměř výhradně v hnilobách na končetinách napadených v předchozí sezóně. Největší počet rakovin, a tedy i nejdůležitějších zdrojů inokula, se vyskytuje na končetinách menších než 38 mm v průměru, zejména v okolí řezů provedených v předchozím roce za účelem odstranění napadených končetin. Během časného jara se v reakci na vyšší teploty a rychlý vývoj pupenů začnou bakterie na okrajích rakoviny rychle množit a vytvářet hustý žlutavý až bílý maz, který se na povrchu kůry vyvíjí až několik týdnů před obdobím květu. Mnoho druhů hmyzu (převážně mušek) je přitahováno výtokem, který následně roznáší bakterie po celém sadu. Jakmile je prvních několik otevřených květů kolonizováno bakterií, opylující hmyz rychle přenáší patogen na další květy, čímž iniciuje další napadení květů. Tyto kolonizované květy podléhají infekci během několika minut po jakémkoli smáčení způsobeném deštěm nebo silnou rosou, pokud jsou průměrné denní teploty rovny nebo vyšší než 16 °C, dokud jsou květní plátky neporušené (květní úbory a mladé plody jsou po opadu plátků odolné). Jakmile dojde k infekci květů, lze očekávat časné příznaky při nahromadění alespoň 57 stupňů denně (DD) vyšších než 13 °C, což může v závislosti na denních teplotách vyžadovat 5 až 30 kalendářních dnů.
S výskytem příznaků květopasa se počet a rozmístění zdrojů inokula v sadu výrazně zvyšuje. Inokulum z těchto zdrojů se dále šíří větrem, deštěm a mnoha náhodnými návštěvníky z řad hmyzu na mladé vrcholky výhonů, čímž se zvyšuje pravděpodobnost vypuknutí plísně výhonů. Nedávný výzkum provedený v Pensylvánii naznačuje, že žír mšic nepřispívá ke vzniku padlí výhonů. Je zapotřebí dalšího výzkumu, aby se zjistilo, zda listonoši hrají roli při výskytu padlí výhonů. K většině infekcí špiček výhonů dochází v období mezi tím, kdy mají výhony přibližně devět až deset listů a nasazením terminálního pupenu, kdy jsou k dispozici zdroje inokula a hmyzí přenašeči a kdy denní teploty dosahují v průměru 16 °C nebo více.
V letech, kdy nedochází k infekci květů, jsou hlavním zdrojem inokula pro fázi napadení výhonů přezimující hálky a zejména mladé výhonky v blízkosti těchto hálek, které se infikují, protože se do nich bakterie dostávají systémově z okrajů hálek. Takovéto systémové infekce výhonů, nazývané rakovina výhonů, jsou zřejmě iniciovány asi 111 DD nad 13 °C po zelené špičce, ačkoli viditelné příznaky se mohou projevit až po nahromadění nejméně 157 DD nad 13 °C po zelené špičce. Při absenci infekce květů je rozvoj infekce padlím výhonů často lokalizován kolem oblastí s přezimujícími rakovinami.
Modelová květní plíseň
Model vyžaduje, aby uživatel rozpoznal specifické a neustále se měnící místní události a aspekty svého sadu, které mohou zvýšit nebo snížit riziko požáru v porovnání s jinými sady v regionu. Model vyžaduje, aby uživatel předpokládal, že riziko infekce ohnivkou existuje vždy, když jsou na stromech přítomny květy, zejména v období opadu okvětních lístků a po odkvětu, kdy na mnoha odrůdách jabloní a hrušní mohou zůstat rozptýlené květy. Uživatel modelu je vyzván, aby pečlivě posoudil situaci na svém konkrétním stanovišti a zahájil kontrolní opatření, pokud jsou přítomny květy, úroveň rizika je "vysoká" nebo "extrémní" a je pravděpodobné, že někdy během následujících 24 hodin dojde k namočení květů.
Struktura modelu: Teploty a vlhkost: Klíčovým procesem ohnivé plísně, který je třeba modelovat, je možnost růstu bakterií na stoncích květů jabloní a hrušní. Tento růst je závislý na teplotě, takže spolehlivá předpověď rizika infekce vyžaduje použití metody měření, která co nejpřesněji odráží růst bakterií. Erwinia amylovora kolonie. Hlavní neshoda mezi modeláři panuje v tom, jak by to mělo být provedeno.
Model Cougarblight odhaduje rychlost růstu bakterií s počtem stupňů v hodinách na základě specifické křivky rychlosti růstu. Tato růstová křivka je založena na rychlosti růstu E. amylovora bakterie v laboratorních testech. Hodnoty hodinových stupňů se sčítají za každou hodinu dne, kdy teplota přesáhne 15 °C. Hodinové hodnoty se zvyšují s nárůstem teplot od 15 °C do 29 °C, při vyšších teplotách klesají a při teplotách nad 40 °C dosahují nulové hodnoty za hodinu.
V aplikaci FieldClimate.com jsou oba modely zobrazeny na stejném obrázku. Model Fireblight DIV vychází z modelu Courgarblight a model Blossomblight vychází z modelu Maryblight.
Model puma nákaza
Výskyt květní plísně je jedním z nejspornějších aspektů epidemií ohnivé plísně. Model byl vytvořen na základě předpokladu, že existuje hojnost inokula a že aby došlo k infekci květů, musí být postupně splněny čtyři přísné podmínky. Tyto podmínky jsou následující:
1) květ musí být otevřený s neporušenými stigmaty a okvětními lístky, stigmata musí být obnažená pro kolonizaci, květy v opadu okvětních lístků jsou odolné;
2) Kumulace alespoň 110 hodin s teplotou > 18,3 °C během posledních 66 dnů s teplotou > 4,4 °C definuje epifytický infekční potenciál nejstaršího otevřeného, a tedy nejvíce kolonizovaného květu v sadu.
3) zvlhčení v podobě rosy nebo 0,2 mm deště nebo 2,5 mm deště v předchozím dni umožňuje přesun bakterií z kolonizovaných stigmat do nektarthod.
4) Průměrná denní teplota >= 15,6 °C: To může mít vliv na rychlost migrace bakterií do nektarthod i na množení bakterií, které je nutné pro vznik infekce.
Grafické znázornění modelů
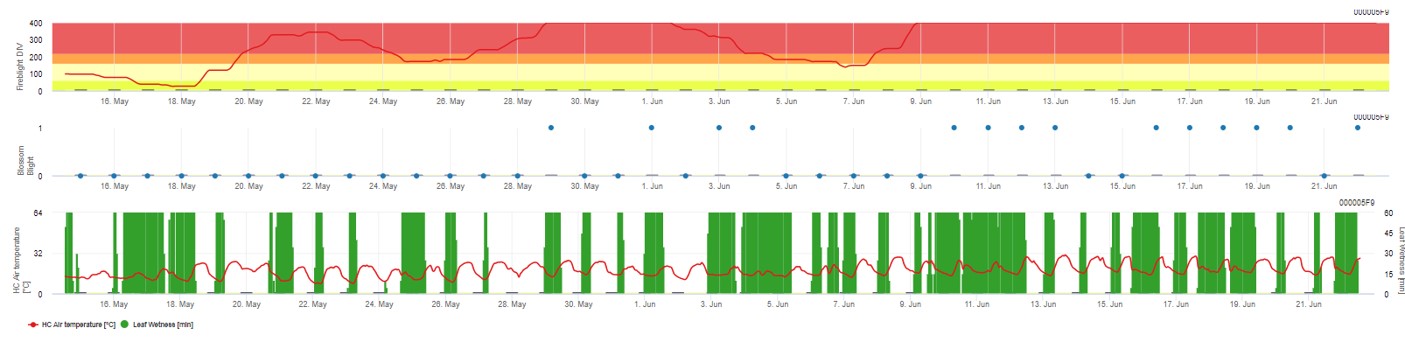
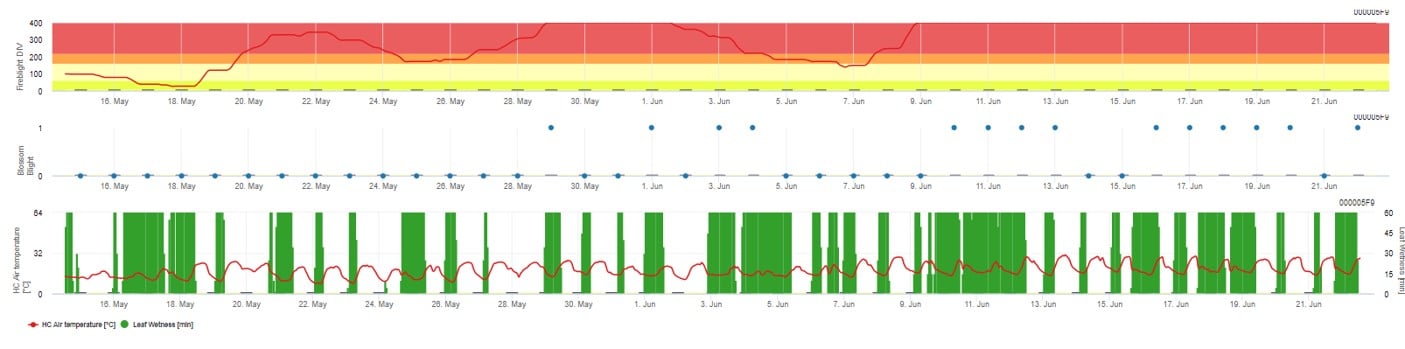
Oba modely jsou zobrazeny na stejné stránce. Grafické znázornění modelů požáru je nejvhodnější používat s hodnotami agregovanými na denní bázi. V grafu jsou zobrazeny hodnoty klimatu související s touto chorobou. Výsledek pumy s názvem Fire Blight DIV a výsledky Blossom Blight. Pro interpretaci výsledků Cougar Blight je graf podložen 5 různými barvami. Rozložení těchto barev je provedeno na základě nastavení na první historii výskytu puchýřky v sadu (viz tlačítko vpravo nahoře) . Těchto 5 barev označuje třídu rizika pro hodnoty DIV.
Interpretace hodnot (v závislosti na výskytu požáru v posledních letech, základní nastavení):
Aktivní rakovina v blízkosti
Nastavení historie požáru nastavte pomocí modrého spodního tlačítka "Nastavení" vpravo nahoře:
Zobrazení výsledků modelu Fire Blight pomocí 1) Modell Cougar Blight (nazvaný Fireblight DIV na základě historie a barev pozadí) a 2) Modell Blossom Blight.
| Potenciál výskytu patogenů | Nízká | Mírná | Vysoká | Extrémní |
| Potenciál výskytu patogenů | Nízká | Mírná | Vysoká | Extrémní |
| Žádné požáry v posledních dvou letech | 0-350 | 350-500 | 500-800 | 800+ |
| Požární nákaza v místní oblasti dvě sezóny | 0-300 | 300-500 | 500-750 | 750+ |
| Požární nákaza v místní oblasti dvě sezóny | 0-250 | 250-450 | 450-700 | 700+ |
| Ohnivá nákaza v sadu v loňském roce | 0-200 | 200-350 | 350-500 | 500+ |
Akční práh:
Kontrola se doporučuje, pokud je zjištěno období s vysokým nebo extrémním rizikem infekce. Faktory určující poškození jsou: počet květů, stáří stromu, vitalita a odrůda,... také.
Možnost infekce Blossom Blight je zobrazena sloupcem od 0 do 1 (všechny podmínky jsou tedy splněny nebo ne).
Cílem modelů ohnivé plísně je posoudit pravděpodobnost infekce prostřednictvím Erwinia amyloflora v sadu.
Model Mary Blight, který vyhodnocuje květní plíseň, velmi dobře indikuje infekční situace s vysokým ekonomickým dopadem. Z tohoto důvodu se poměrně často používá k indikaci použití antibiotik proti tomuto patogenu. Puma blight poskytuje informace o riziku infekce ohnivkou do k celkovým možnostem šíření patogenu. Její vážení provedené na základě historie sadu je velmi užitečné, aby nám naznačilo, jak pečlivě máme kontrolovat sady na příznaky ohnivé plísně i v situacích, kdy Mary Blight nebude indikovat infekci.
Biologie můry tesaříka
Na stránkách C. pomonella přečkává zimu jako zralá larva stočená do kokonu, který se nachází na chráněných místech na stromě (štěrbiny nebo praskliny v kůře), což je preferované místo, nebo u paty stromu v podestýlce. V moderních jabloňových sadech s vysokou hustotou je na stromech kvůli hladké kůře jen velmi málo míst pro zakuklení. Kukly se tvoří v kokonu, který spřádá dospělá larva.
Dospělé stadium se vylíhne z kukly a po krátké době zpevnění exoskeletu vletí na strom. Většina literatury uvádí, že aktivita můry probíhá v horní třetině koruny stromu. Dospělci jsou aktivní za soumraku po dobu několika hodin, přičemž průměrný práh aktivity se pohybuje mezi 13 °C a 15 °C. K páření obvykle dochází na hostiteli a bezprostředně poté dochází ke kladení vajíček. Za teplých večerů je možné, že se samice můry vylíhne, spáří a naklade vajíčka během dvou dnů. A C. pomonella v ideálních podmínkách může žít více než 20 dní, na polích však pravděpodobně přežije mnohem kratší dobu, zejména v horkém létě. V laboratorních podmínkách naklade samička až 100 vajíček, ale skutečná plodnost v terénu je pravděpodobně nižší a většina z nich je nakladena v prvních dnech po objevení a páření.
Vajíčka jsou kladena jednotlivě na horní stranu listu, na větvičky nebo na plody. V první generaci jsou vajíčka kladena pouze na listy, a to přibližně do dvou týdnů po nasazení plodů, kdy se ochmýření plodů změní. Ve druhé generaci je asi 65% vajíček kladeno na listy, ale ty jsou obvykle velmi blízko plodu. Vajíčka jsou při prvním kladení krémově bílá, v polovině vývoje mají červený kroužek a v době blízké líhnutí je vidět černá hlavička tobolky.
Čerstvě vylíhlá larva bloudí, dokud nenajde plod. Prokousne slupku a několik dní se živí dužinou jablka pod slupkou. Při prokousávání slupky a žíru pod ní larva vytlačuje výkaly z místa vstupu. Kolem vniknutí se často vytvoří červený prstenec, zejména u mladých plodů. Poté, co larva přejde do dalšího stádia, zavrtá se směrem k jádřinci, kde se až do dospělosti živí semeny. Když larva téměř dospěje, vytvoří tunel od jádřince k vnější straně jablka a otvor ucpe výkaly a hedvábím. Když je larva připravena vytvořit zámotek, opustí plod a na hedvábném vlákně se pohybuje po stromě nebo spadne na zem a najde si chráněné místo.
Model pro mol Codlingův
Náš model je založen na kumulaci efektivních teplot mezi >= 10°C a <=32°C. Na severní polokouli začíná tato akumulace prvním lednem. Na jižní polokouli začíná prvním červencem. Každé měření kumulujeme v jeho ekvivalentu pro daný den (viz graf níže). Pro vývoj hmyzu se používají následující prahové hodnoty:
*Kompletní generace= 650
| *Skladení vajec až líhnutí vajec | = 88 |
| *První stádium larev | = 60 |
| *Druhé stadium larev | = 65 |
| *Třetí stádium larev | = 55 |
| *Stádium larvy | = 45 |
| *Páté stadium larev a kukly | = 335 |
Graf vytvořený modelem zobrazuje první výskyt a dobu trvání larvového instaru 1 až 5 a přítomnost dospělců v 5 úrovních.
| *Úroveň 1: | Letová aktivita, jiná než snáška vajec |
| *Úroveň 2: | Letová aktivita s možným kladením vajec. |
| *Úroveň 3: | Letová aktivita s určitou snůškou vajec. |
| *Úroveň 4: | Letová aktivita s přiměřenou až silnou snůškou vajec. |
| *Úroveň 5: | Letová aktivita se silnou snůškou vajec. |
Prezentace stádií dospělců můry tesaříka v pěti různých úrovních (první graf), jakož i kumulovaných stupňových dnů (červená čára, druhý graf) pro vývoj dospělců a modře znázorněných larválních stádií (třetí graf).
V jabloňových sadech se vyskytuje několik druhů mšic. Jedná se o mšici zelenou, mšici ovesnou nebo mšici jabloňovou a mšici růžovku. V soukromých zahradách má určitý význam především růžová mšice jabloňová. Tato mšice vede při napadení k těžkým hormonálním změnám na koncích výhonů. Listy se zkroutí a výkvět se zkrátí. Později listy žloutnou a odumírají. Tato mšice napadá především mladé výhony. Mšice růžovková na jabloni může způsobit vážné škody již při malém množství. Zatímco mšice zelená jabloně a mšice jabloňová se mohou vyskytovat ve větším množství, aniž by vedly k hospodářským škodám. Mšici jabloňovou je třeba hodnotit jako hospodářsky nevýznamnou vůbec.
Mšice jabloňová růžová
Jedná se o starý evropský druh, který byl do Spojených států dovezen kolem roku 1870. Koncem 19. století se stal významným škůdcem jablek. Aby se tomuto druhu dařilo, musí mít hojnost své letní hostitelské rostliny, jitrocele úzkolistého. Mezi zavlečením a rozšířením mšice růžkaté a jitrocele úzkolistého existuje pozoruhodná paralela. Tato mšice se vyskytuje ve všech ovocnářských oblastech na celém světě.
Životní fáze
Vejce: Vejce je oválné a na straně u kůry mírně zploštělé. Délka se pohybuje od 0,49 do 0,56 mm. Při prvním kladení je jasně žluté a pokryté lepkavou hmotou, která s věkem tvrdne. Barva se postupně mění na zelenožlutou a nakonec na leskle černou. Doba potřebná k této změně barvy se v běžných venkovních podmínkách pohybuje od přibližně devíti dnů do více než dvou týdnů.
Nymfa: Z vajíček se vylíhnou samci bez křídel, kteří jsou živorodí. Existuje pět instarů. Posledním je zralá stonková matka, která krátce po čtvrtém instaru začíná partenogeneticky produkovat živá mláďata. Denně vyprodukuje v průměru pět až šest mláďat. Nymfy druhé generace, které jsou všechny samičí, dosahují dospělosti za dva až tři týdny. Převážná většina nymf se začíná rozmnožovat na jabloni, i když několika z nich se mohou vyvinout křídla a migrovat na plantáž.
Třetí generace se vyrábí v červnu a na začátku července. Většině této generace se vyvinou křídla a migrují na úzkolisté ploštice. V některých ročních obdobích bezkřídlé samičky třetí generace vytvářejí čtvrtou generaci na jabloni. V posledních letech bylo v některých oblastech pozorováno, že škodlivé populace mšice růžkaté přetrvávají v sadech až do poloviny léta nebo i později. Není známo, zda je tato změna způsobena selekcí, změnou habitu nebo nedostatkem potřeby alternativního hostitele.
Dospělí: Dospělí jedinci se značně liší barevnými znaky. Celkové zbarvení je růžově hnědé, s narůžovělým nádechem způsobeným práškovitým povlakem. Někteří starší dospělci jsou fialoví, zatímco mladší dospělci jsou rozhodně červenorůžoví.
Rozsah hostitele
Nejoblíbenějším hostitelem je jabloň, ale mšice se živí také hrušní a hlohem. Obzvláště náchylné jsou odrůdy jablek Cortland, Golden Delicious, Rhode Island Greening a Ida Red. Mšice se vyskytuje ve všech ovocnářských oblastech Spojených států a Kanady.
Zranění nebo poškození: Žír mšice růžové často způsobuje kroucení listů jabloní, které začíná při opadu okvětních lístků. Tyto listy se později mohou zbarvit do jasně červené barvy. Krmení na listech v okolí plodů má často za následek shlukování, zakrňování a deformaci vyvíjejících se plodů. Tyto abnormality se s vývojem plodů zhoršují a nakonec mohou způsobit, že plody nejsou prodejné.
Velké populace mšic mohou produkovat velké množství medovice jako odpad z mízy, kterou se živí. Medovice vylučovaná na plody slouží jako živná půda pro růst plísně sazí, která ovlivňuje povrchovou úpravu jablek. Toxiny ve slinách mšic slouží také jako "stopka", která brání abscesu plodů (přirozenému oddělení od stromu) při běžné sklizni.
Životní styl: Mšice přečkává zimu ve stadiu vajíčka. K líhnutí dochází brzy na jaře, asi o týden až 10 dní později než u mšice obilní a přibližně ve stejnou dobu jako u mšice jabloňové. Vajíčka se líhnou, když se na jaře začnou otevírat pupeny, tedy po dobu dvou týdnů. Jakmile se mláďata vylíhnou, vyhledávají otevírající se pupeny jabloní; zdá se, že dávají přednost ovocným pupenům. Živí se na vnější straně listových a ovocných pupenů, dokud se listy nezačnou rozvíjet. Poté se dostanou dovnitř trsů a začnou sát šťávu ze stopek a nově vytvořených plodů.
Jejich žír způsobuje zkroucení listů, což mšice chrání před postřiky a některými nepřáteli. Silné kroucení listů způsobené tímto druhem je pravděpodobně nejcharakterističtějším znakem jeho činnosti. Jediná stonková matka umístěná na spodní straně listu v blízkosti středního žebra způsobí pevné svinutí listu. Stačí jen několik stonkových matek, aby způsobily silné stočení všech listů v okolí otevírajícího se květního pupenu, což poskytuje ideální ochranu rychle se vyvíjejícím mšicím. Stonkové matečníky dosahují dospělosti v době, kdy jabloně začínají kvést.
Dospělé kmenové matky jsou velmi neaktivní. Usazují se, krmí se a rychle plodí mláďata. Při rozdělení rychle odstraňují zobáky z rostlinných pletiv a hledají jiné místo, kde by mohly pokračovat. Stonkové matky dospívají asi dva týdny po vylíhnutí. Délka této doby závisí do značné míry na povětrnostních podmínkách. Produkce mláďat obvykle začíná dva až tři dny po posledním líhnutí a pokračuje bez přerušení déle než měsíc.
Celková produkce jedné samice je v průměru asi 185 kusů. Období rozmnožování trvá obvykle od začátku května do června. Obvykle je maximum reprodukční aktivity kolem posledního týdne května a prvního týdne června, kdy se začínají zakládat mladé plody a začíná aktivní růst. Mšice jabloňová růžová se vyskytuje jen zřídka a napadá mladé a rychle rostoucí výhony. Omezuje se na listy, květní stopky a mladé plody.
Jedním z charakteristických znaků tohoto druhu je shlukování mláďat kolem matky. Každá jednotlivá kmenová matka nebo skupina matek má kolem sebe stovky mláďat. Napadené listy mohou být brzy pokryty - v některých případech více než jednou vrstvou mšic. Tento zvyk shlukování brzy zničí napadené listy a způsobí nucenou migraci mšic. Mláďata se pohybují aktivně a spěšně, zřejmě se snaží najít vhodnou živnou půdu. V tomto období se často shromažďují na tvořících se plodech nebo napadají nové, šťavnaté rozvíjející se listy.
Druhá generace potřebovala čtyři až 40 dní, aby dosáhla dospělosti a vyvedla mláďata. Většinu druhé generace tvoří samice bez křídel. Průměrná celková produkce každého jedince ve druhé generaci asi 119. Zvyky a aktivity třetí generace se neliší od zvyků a aktivit druhé generace. Mšice se shromažďují v obrovském množství na spodní straně listů a způsobují silné kroucení. Napadají také nasazující a vyvíjející se plody a způsobují charakteristická poranění.
Většina této třetí generace získává křídla a stěhuje se na úzkolistý jitrocel, letní hostitelskou rostlinu. Může se vytvořit čtvrtá generace. Všechny tyto generace vytvářejí křídla a migrují na úzkolistý jitrocel. Po posledním vylíhnutí jsou okřídlení dospělci velmi křehcí a neaktivní. Dva až tři dny zůstávají vylučováni ve stočených listech, než se vydají na migrační let. Těsně před odletem jsou velmi aktivní a nervózní, pobíhají nebo pohybují křídly nahoru a dolů v očekávání letu.
Mšice zelená jabloně
Mšice zelená Aphis pomi (DeGeer) je rozšířená ve všech oblastech pěstování jablek. Na severní polokouli se poprvé objevuje v jabloňových sadech koncem května až začátkem června. Hmyz saje šťávu z listů na vodních klíčcích a šťavnatých terminálních porostech. Mšice zelená jabloně se obvykle vyskytuje v blízkosti hlavních žilek na spodní straně listu. Mšice zelená jabloně napadá také hrušeň, hloh, kdouloň, krabí jablko a jírovec.
Popis mšice zelné
Vajíčka mšice zelené jsou oválná a leskle černá. Nymfy a dospělci jsou tmavě nebo světle zelení. Mšice zelená jabloně má zelenou hlavu, stejnoměrně zelené nebo žlutozelené bříško, černé nohy a dlouhé černé rohoviny - "ocasní roury" (na zadním konci hmyzu). Dospělé samičky jsou tmavě zelené až černé a okřídlené. V létě tyto samičky neboli alaty rodí partenogenně živá mláďata.
Biologie mšice zelné
Mšice zelená přezimuje jako vajíčko na odnožích, na bázi pupenů a na listových jizvách terminálních výhonů. Vajíčka se obvykle líhnou v době, kdy praskají pupeny a rozvíjejí se první listy. Nymfy se okamžitě začínají živit na vyvíjejících se listech. Zpočátku se vyskytují na terminálním výhonu a později se přesouvají na starší listy trsu. Dospělé samičky mšic vytvářejí bezkřídlé samičky během dvou týdnů, přičemž každá samička je schopna vyprodukovat 50-100 živých potomků. Mladé mšice se vyvíjejí sedm až deset dní. Populace mšice zelné se na jabloni obvykle vytvářejí pomalu brzy na jaře (kvetení, okvětní lístky, podzim) a rychleji s rostoucími průměrnými denními teplotami. Během léta se vytvářejí okřídlené mšice neboli alaty a bezkřídlé mšice. Mšice zelená jabloně jsou nejpočetnější v červenci a na začátku srpna. V počátečních fázích napadení je více nymf než dospělců alat. S růstem populace se podíl alat zvyšuje. Kolonie mšice zelné se mohou za příznivých podmínek zdvojnásobit během jednoho týdne. Pokud mšice nemají šťavnatý nový přírůstek a musí se živit staršími listy, může počet vyprodukovaných mláďat klesnout až o 50%. Pokud jsou denní teploty 30-32 °C a vyšší, samičky se špatně rozmnožují. Pokud jsou teploty vysoké po několik dní, mšice hynou. Během léta se vyvíjí několik generací mšice zelené jabloně a okřídlené formy se rozptýlí po celém sadu. Někdy se stává, že mšice zelená a mšice růžová, Dysaphis plantaginea Pars., mají kolonie na stejném listu. Růžová mšice jabloňová je růžová nebo fialová. Na podzim se vyvíjejí a páří bezkřídlí samci a samičky, které kladou vajíčka na zimu. Největší množství vajíček je ve vzdálenosti 15-20 cm od špiček větviček. Vajíčka jsou jen zřídka na velkých lešenářských větvích nebo na kmenech jabloní.
Poškození
Mšice zelená vysává šťávu z listů. Silné napadení snižuje vitalitu a růst výhonů. To se týká zejména školek a mladých, nerodících sadů. Hmyz může snižovat velikost pupenů a délku mezičlánků a způsobovat kroucení listů. Žír mšic může také stimulovat boční růst větví, který může ovlivnit tvar stromu. Kadeřavost listů a oslabené terminály jsou náchylné k zimnímu poškození. Medovice produkovaná mšicemi může stékat na plody a umožnit růst plísní. Houba kazí plody a snižuje jejich tržní hodnotu. Při silném napadení se mšice zelená může živit nezralými jablky a způsobit jejich rzivost.
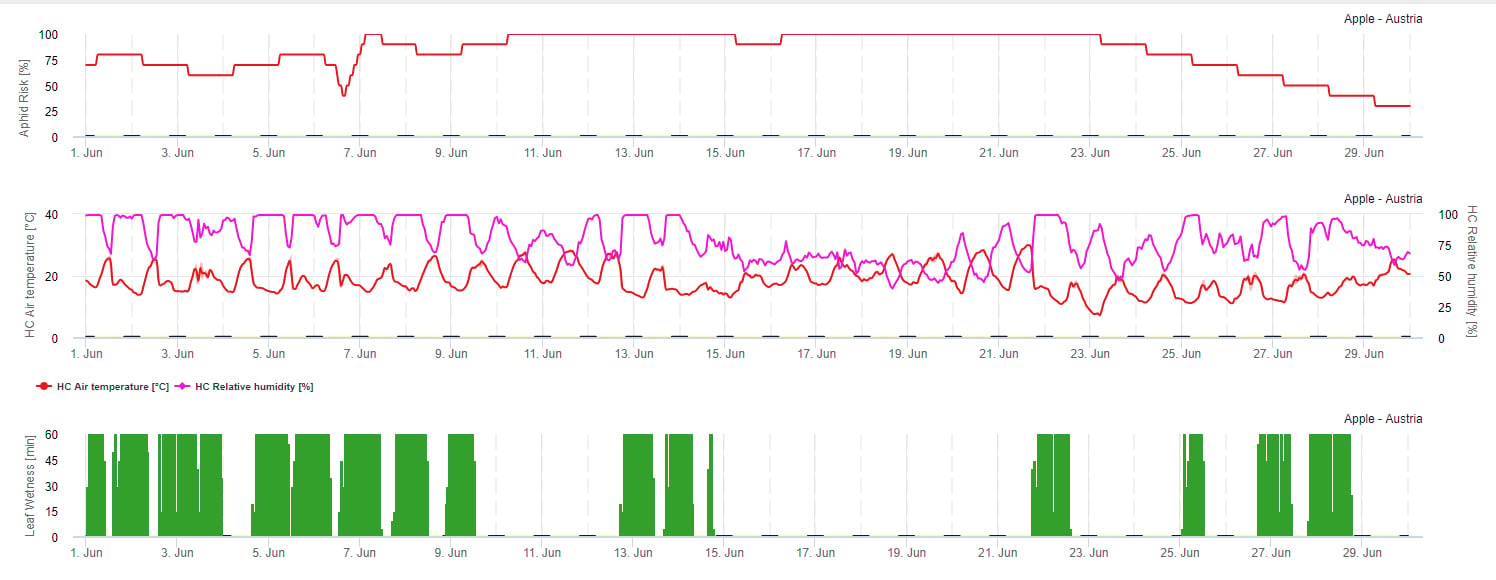
Model rizika šíření
Na šíření mšic má vliv především teplota. Čím více hodin se blíží optimální teplotě (20-32 °C) a je k dispozici zdroj potravy, tím je rychlost množení vyšší. Zatímco vajíčka a mladé instary jsou citlivé vůči suchému vzduchu. Nízká relativní vlhkost vzduchu může populaci mšic také poškodit (pod 30% r.h.). Kromě insekticidů jsou mšice potlačovány predátory a parazitickými houbami. Parazitické houby jsou závislé na vlhkosti. Většina těchto druhů jako např. Beauvaria bassiana jsou zvýhodněny volnou vlhkostí.
Riziko zvyšujeme, když je optimální teplota mezi 20 a 32 °C a relativní vlhkost mezi 30 a 95% r. v., a snižujeme, když je příliš vlhko (vlhkost listů, déšť nebo relativní vlhkost nad 95%), příliš chladno (teploty nižší než 20 °C) nebo příliš horko (teploty nad 32 °C), vlhké noci (vlhkost listů v nočních hodinách).